
Venturing into Qualitative Research: A Practical Guide to Getting Started
Scholarship and Practice of Undergraduate Research Journal
- Journal Home
- Submissions
In this commentary, we offer an introduction to qualitative research. Our goal is to provide guidance so that others can avoid common missteps and benefit from our lessons learned. We explain what qualitative data and research are, the value of qualitative research, and features that make qualitative research excellent, as well as how qualitative data can be collected and used to study undergraduate research. Our advice and recommendations are targeted at researchers who, like us, were first trained in fields with tendencies to overlook or underestimate qualitative research and its contributions. We share examples from our own and others’ research related to undergraduate research settings. We provide a table of resources researchers may find useful as they continue to learn about and conduct qualitative studies.
Introduction
We both started our scholarly journeys as biologists. As we trained, we both grew interested in researching undergraduate education and we transitioned to doing education research. We quickly came to realize that our training in experimental approaches and quantitative methods was woefully insufficient to study the diversity of ways students think, believe, value, feel, behave, and change in a variety of learning environments and educational systems.
For instance, there are established ways to quantify some educational variables, but not others. In addition, there may be phenomena at play that we haven’t thought of or that might be counterintuitive, which could lead us to quantify things that end up being irrelevant or meaningless. Herein lies the power of qualitative research. Qualitative research generates new knowledge by enabling rich, multifaceted descriptions of phenomena of interest, known as constructs (i.e., latent, unobservable variables), and producing possible explanations of how phenomena are occurring (i.e., mechanisms or relationships between constructs in different contexts and situations with different individuals and groups).
In this essay, we aim to offer an approachable explanation of qualitative research, including the types of questions that qualitative research is suited to address, the characteristics of robust qualitative research, and guidance on how to get started. We use examples from our own and others’ research to illustrate our explanations, and we cite references where readers can learn more. We expect Scholarship and Practice of Undergraduate Research (SPUR) readers from disciplines with a tradition of qualitative research might question why we would write this piece and what makes us qualified to do so. There are many scholars with much more qualitative research expertise than we have. Yet, we think we can offer a unique perspective to SPUR readers who are new to qualitative research or coming from disciplines where qualitative research is unfamiliar or undervalued. We have both designed, conducted, and published qualitative research in the context of undergraduate education and research experiences. We draw upon this experience in the recommendations we offer here.
Doing qualitative research involves acknowledging your “positionality,” or how your own background, lived experiences, and philosophical understandings of research influence how you approach and interpret the work (e.g., Hampton, Reeping, and Ozkan 2021; Holmes and Darwin 2020). Our positionalities have influenced our approach to this article and qualitative research generally. I (MAP) first learned about qualitative research from my undergraduate academic adviser. She invited me to help her implement and evaluate a capstone course in which groups of microbiology undergraduates engaged in a semester-long research project to address problems faced by community organizations (Watson, Willford, and Pfeifer 2018). At the time, I wasn’t aware of the long-standing history of qualitative research or its different forms and approaches. I just knew that reading quote data helped me understand human experiences in a way that survey numbers did not. Since my introduction to qualitative research, I’ve been fortunate to receive formal training. I consider my most valuable lessons about qualitative research to be through the practical experience of doing qualitative research and being mentored by qualitative researchers.
When I (ELD) first learned about qualitative research, I thought it meant words – perhaps collected through surveys, focus groups, interviews, or class recordings. I thought qualitative research would be easy – it was just words after all, and I had been using words almost my whole life. I assumed if I collected some words and summarized what I thought they meant (think word cloud), I would be doing qualitative research. As we will elaborate here, this is a limited view of what qualitative research is and what qualitative research can accomplish. When I began presenting qualitative research, I found it helpful to draw analogies to qualitative studies in natural science and medical disciplines. For instance, in the field of biology, the invention of technologies (e.g., lenses, microscopes) allowed for detailed observation and rich descriptions of cells (i.e., qualitative research) that led to the development of cell theory, the establishment of the field of cell biology, and quantitative research on cell structure, function, and dysfunction. In my own field of neuroscience, Henry Moliason, known as HM, was the focus of qualitative case study because he lost the ability to form new long-term memories due to a surgical treatment for severe epilepsy. Rich (i.e., comprehensive and detailed) description of Mr. Moliason’s memory impairment was the basis for hippocampal function being proposed as the main mechanism through which memories are formed. These examples of “non-numbery” research that produce influential descriptions and testable mechanisms helped me recognize the potential value and impact of qualitative research.
Types of Qualitative Research Questions
Qualitative research is useful for addressing two main types of questions: descriptive and mechanistic. Descriptive questions ask what is happening, for whom, and in what circumstances. Mechanistic questions ask how a phenomenon of interest happening. Here we explain each type of question and highlight some example studies conducted in the context of undergraduate research.
Descriptive Questions
Descriptive research seeks to elucidate details that enhance our overall understanding of a particular phenomenon—it answers questions about what a phenomenon is, including its defining features (i.e., dimensions) and what makes it distinct from other phenomena (Loeb et al. 2017). Descriptive research can also reveal who experiences the phenomenon, as well as when and where a phenomenon occurs (Loeb et al. 2017). Details like these serve as a starting point for future research, policy development, and enhanced practice. For instance, Hunter, Laursen, and Seymour (2007) carried out a qualitative study that identified and described the benefits of undergraduate research from the perspectives of both students and faculty. This work prompted calls for expansion of undergraduate research nationally and led to numerous quantitative studies (Gentile, Brenner, and Stephens 2017). Among these were quantitative studies from our group on the influences of research mentors on undergraduate researchers (Aikens et al. 2016, 2017; Joshi, Aikens, and Dolan 2019). Although these studies were framed to identify beneficial outcomes, we observed that undergraduates who had less favorable experiences with mentors were opting not to participate in our studies. Given this observation and the dearth of research on negative experiences in undergraduate research, we carried out a descriptive qualitative study of the dimensions (i.e., the what) of negative mentoring—that is, problematic or ineffective mentoring—in undergraduate life science research (Limeri et al. 2019). This study revealed that negative mentoring in undergraduate research included the absence of support from mentors and actively harmful mentor behaviors. These results served as the basis for practical guidance on how to curtail negative mentoring and its effects and for ongoing quantitative research. We use this study as the basis for the extended examples highlighted in Table 1.
Descriptive research is also suited to investigating the experiences of groups that are marginalized or minoritized in higher education. These studies offer insights into student experiences that may be otherwise overlooked or masked in larger quantitative studies (Vaccaro et al. 2015). For example, descriptive qualitative research shed light on how Black women in undergraduate and graduate STEM programs recognized and responded to structural racism, sexism, and race-gender bias. This research identified how high-achieving Black STEM students experienced racial battle fatigue and offered program-level suggestions for how to better support Black students (McGee and Bentley 2017). Descriptive qualitative research of deaf students involved in undergraduate research revealed that lack of awareness of Deaf culture of research mentors as well as lack of communication hindered students’ research experiences (Majocha et al. 2018). This research led to recommendations for research programs, research mentors, and students themselves. Another descriptive qualitative study showed how Latine students’ science identity changed over time when involved in an undergraduate research program (Vasquez-Salgado et al. 2023). Specifically, Vasguez-Salgado and colleagues identified patterns in students’ science identity through three waves of data collection spanning 18 months. Students’ identities showed consistent or fast achievement of feeling like a scientist, gradual achievement of feeling like a scientist, achievement adjustment of feeling like a scientist at one point and less so later in the program, or never feeling like a scientist. Together, these and other studies have generated knowledge that raises questions for future research and informs our collective efforts to make undergraduate research more accessible and inclusive.
Mechanistic Questions
Mechanistic qualitative research aims to address questions of how or why a phenomenon occurs. In the context of undergraduate research, an investigator may seek to understand how or why a particular practice or program design affects students. Recently, we conducted a mechanistic qualitative study that aimed, in part, to understand how early career researchers (undergraduate, postbaccalaureate, and graduate students) conceptualized their science identity (Pfeifer et al. 2023). Previous research theorized that someone is more likely to identify as a scientist if they are interested in science, believe they are competent in and can perform science, and feel recognized by others for their scientific aptitude or accomplishments (Carlone and Johnson 2007; Hazari et al. 2010; Potvin and Hazari 2013). However, this theory is somewhat limited in that it does not fully explain how context affects science identity or how science identity evolves, especially as researchers advance in their scientific training (Hazari et al. 2020; Kim and Sinatra 2018). To address this, we integrated science identity theory with research on professional identity development to design our study (Pratt, Rockmann, and Kaufmann 2006). We analyzed data from two national samples, including open-ended survey responses from 548 undergraduates engaged in research training and interview data from 30 early career researchers in the natural sciences. We found that they conceptualized science identity as a continuum that encompassed being a science student, being a science researcher, and being a career researcher. How students saw their science identity depended on how they viewed the purpose of their daily research, the level of intellectual responsibility they have for their research, and the extent of their autonomy in their research. We consider these findings to be hypotheses that can be tested quantitatively to better understand science identity dynamics in research training contexts. By asking this mechanistic question about science identity, we sought to add to and refine existing theory.
Key Attributes of Qualitative Research
For any type of research to be meaningful, it must possess some degree of rigor—what qualitative researchers call trustworthiness (Morse et al. 2002; Yilmaz 2013). Qualitative research is more trustworthy if it is characterized by credibility, transferability, dependability, and confirmability (Creswell and Poth 2016; Lincoln and Guba 1985). For instance, like accuracy and precision in quantitative research, do qualitative findings reflect what is being studied and are the interpretations true to the data (credibility)? Similar to reproducibility in quantitative research, how can qualitative research findings be applied to similar contexts (transferability)? Like validity in quantitative research, to what degree are the framing, methods, and findings of qualitative research appropriate given the aims (dependability)? Similar to the idea of replicability in quantitative research, if the same analytic tools were applied to the same data set could similar findings be reached by someone outside the original research team (confirmability)? The exact dimensions of trustworthiness, how trustworthiness manifests in the research process, the best ways to achieve trustworthiness, and how to talk about trustworthiness in research products are the subject of ongoing and often-spirited debate (e.g., Gioia et al. 2022; Mays and Pope 2020; Morse et al. 2002; Ritchie et al. 2013; Tracy 2010; Welch 2018; Yadav 2022). Central to these dialogues is the fact that qualitative research is composed of different philosophical approaches that emerged and evolved from diverse social science fields (Creswell and Poth 2016; Ritchie et al. 2013). Identifying universally agreed-upon criteria and the means to achieve these criteria is complex.
In our own work, we have found Tracy’s (2010) eight criteria for excellent qualitative research particularly useful. These criteria have helped us design studies, make decisions during the course of research, and articulate in our papers how our research seeks to achieve trustworthiness (e.g., Pfeifer, Cordero, and Stanton 2023). The full list of criteria is: worthy topic, rich rigor, sincerity, credibility, resonance, significant contribution, ethical conduct, and meaningful coherence (Tracy 2010). These criteria borrow from and build on the presented concepts of credibility, transferability, dependability, and confirmability. In our view, these criteria are presented and described in a way that makes sense to us and fits our approach to research. Here we highlight two criteria that may be particularly relevant if you are new to qualitative research.
Worthy Topics
As scholars familiar with undergraduate research and scholarly inquiry, SPUR readers are well-positioned to design studies that address research questions that are significant and timely in the context of undergraduate research. The first step in doing qualitative research (or any research) is to figure out what you want to study. You’ll want to select a topic that you find interesting, relevant, or otherwise compelling so you are motivated to spend time and effort investigating it. One way to find a topic is to notice what is happening in your environment and your work. What are you observing about undergraduate research? Something about students who participate (or not)? Something about colleagues who work with undergraduate researchers (or not)? Something about the design, implementation, or outcomes of the research experience? Something about the programmatic or institutional context? For a topic to be worthy of research, it should be interesting to you and to others. Consider sharing your observations with a few critical friends (i.e., trusted colleagues who will give you honest feedback) about whether they find your observations interesting or worth your time and energy to explore.
Like other human research, qualitative studies must adhere to basic ethical principles of respect for persons, beneficence, and justice (National Commission for the Protection of Human Subjects 1978). Respect for persons means treating all people as autonomous and protecting individuals with diminished autonomy (e.g., students whom we teach and assess). Beneficence involves treating people in an ethical manner, including respecting their decisions, protecting them from harm, and securing their well-being. Justice refers to the balance between benefiting from research and bearing its burdens; in other words, people should be able to benefit from research and should not be expected to bear the burden of research if they cannot benefit. Although it is beyond the scope of this essay to provide guidance on how to adhere to these principles, it is important to recognize that qualitative methods like interviewing can be highly personal and sometimes powerful experiences for both participants (and researchers). Investigators should carefully consider how their participants may be affected by data collection. For example, you may interview or survey participants about a personally difficult or painful experience. Do you then bear responsibility for helping them find support to navigate these difficulties? What if a participant reveals to you a serious mental health issue or physical safety concern? These situations occurred during our negative mentoring studies. We provided information to participants about where they could seek counseling or support for specific issues that can occur with mentors, such as harassment and discrimination.
Certainly not all qualitative data collection brings up these issues, but it can and does happen more frequently than you might expect. Your institutional review board (IRB), collaborators, and critical friends can be helpful resources when planning for and navigating tough scenarios like this. If working with an IRB is new to you, we recommend finding colleagues at your institution who have conducted IRB-reviewed research and asking them for guidance and examples. Some IRBs offer training for individuals new to developing human research protocols, and there are likely to be templates for everything from recruitment letters to consent forms to study information. We have found the process of developing IRB protocols helps refine research questions and study plans. Furthermore, IRB review is needed before you collect data that will be used for your study; IRBs rarely if ever allow for retrospective review and approval. In our experience, these studies are likely to be determined as exempt from IRB review because they involve minimal risk and use standard educational research procedures. However, the IRB is still responsible for making this determination and is a valuable partner for helping investigators navigate sensitive or complex situations that occur in human research.
Getting Started with Qualitative Research
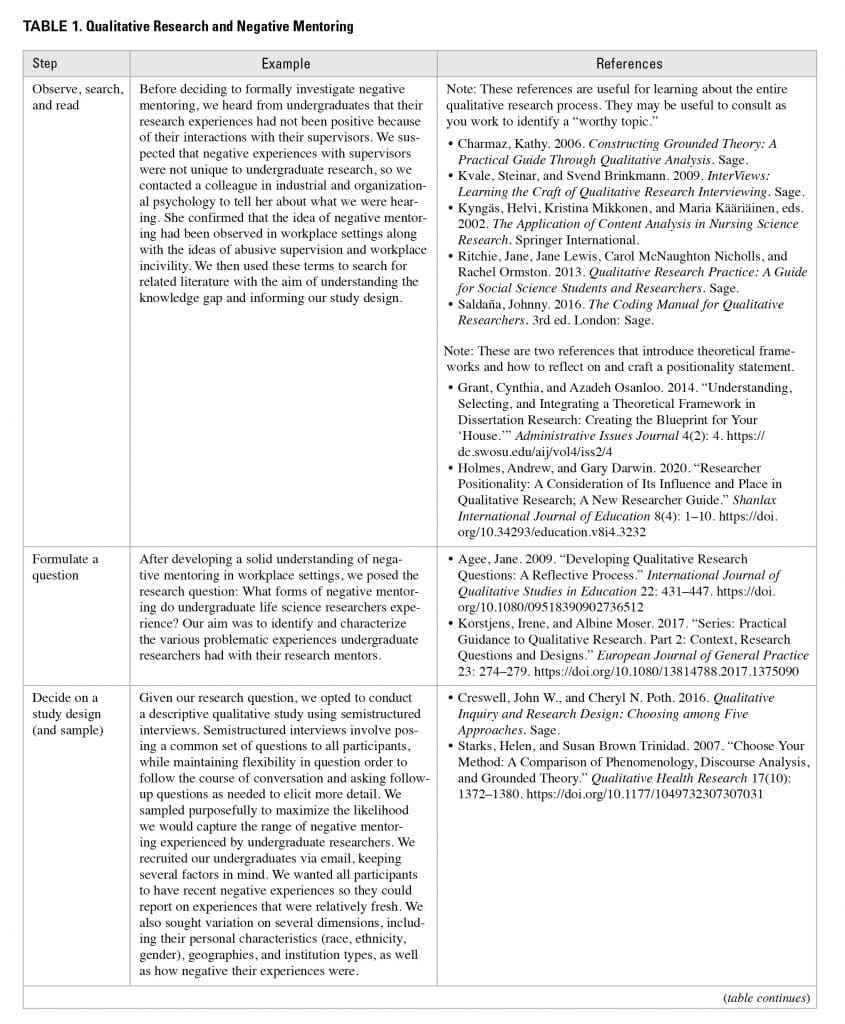
Now that you have a sense of the purposes of qualitative research and what features help to ensure its quality, you are probably wondering how to do it. We want to emphasize that there are entire programs of study, whole courses, and lengthy texts that aim to teach qualitative research. We cannot come close to describing what can be learned from these more substantial resources. With this is mind, we share our own process of carrying out qualitative research as an example that others might find helpful to follow. We outline this “how to” as a series of steps, but qualitative research (like all research) is iterative and dynamic (University of California Museum of Paleontology 2022). Feel free to read through the steps in a linear fashion but then move in non-linear ways through the various steps. Extended discussion of each of these steps with examples from our research on negative mentoring is provided in Table 1 along with an abridged list of our go-to references.
Observe, Search, and Read
For a topic to be worthy of qualitative research (or any research), it should also have the potential to address a knowledge gap. After we identify a “worthy topic,” we try to find as much information about that topic as possible (Dolan 2013). We read, then we keep reading, and then we read some more. This may seem obvious, but we find that investing time reading literature can save us a lot of time designing, conducting, and writing up a study on a phenomenon that is already well known or understood by others and just not (yet) by us. To help us in our searching, we will sometimes reach out to colleagues in related fields to describe the phenomenon we are interested in studying and see if they have terms that they use to describe the phenomenon or theories they think are related. Theory informs our research questions, study designs, analytic approaches, and interpretation and reporting of findings, and enables alignment among all of these elements of research (e.g., Grant and Osanloo 2014; Luft et al. 2022; Spangler and Williams 2019). Theory also serves as a touchstone for connecting our findings to larger bodies of knowledge and communicating these connections in a way that promotes collective understanding of whatever we are investigating.
Formulate a Question
Once you have selected a topic and identified a knowledge gap, consider research questions that, if answered, would address the knowledge gap. Recall that qualitative research is suited to questions that require a descriptive (what) or mechanistic (how) answer.
Decide on a Study Design
Just like quantitative research, qualitative research has characteristic approaches, designs, and methodologies, each of which has affordances and constraints (Creswell and Poth 2016; Merriam 2014; Miles, Huberman, and Saldana 2014). Creswell and Poth provide a valuable resource for learning more about different types of qualitative research study designs, including which designs are suited to address which kinds of research questions. Given the labor intensiveness of qualitative data collection and analysis, it is critical to think carefully about how to recruit and select study participants. What this looks like and who might be appropriate study participants will depend on many factors, including the knowledge gap, research question, study design, and methods. Questions that can be helpful to ask are: Who do I need to study to answer my research question? What should the study participants have in common? In what ways should study participants vary to provide rich, complex, and varied insight into what I am studying? To whom do I want to generalize my findings, keeping in mind the qualitative nature of the work?
Based on the answers to these questions, you may opt for purposeful sampling in which you collect data only from participants who meet the characteristics you decide upon given the aims of your study. In this case, you will likely send a screening survey to potential participants to determine what their characteristics of interest are, which will help you decide if you will invite them for further data collection or not. A purposeful sample contrasts with a convenience sample where essentially any person who agrees to participate in the study will be selected for further data collection.
Collect and Analyze Data Systematically
Qualitative data can be collected in a variety of ways, including surveys, interviews, and focus groups, as well as audio and video recordings of learning experiences such as class sessions. To decide which method(s) to use for data collection, it is helpful to consider what you aim to learn from study participants. Surveys tend to be easier to distribute to a larger sample, but may elicit shorter or shallower responses, which are challenging to interpret because there is less information (i.e., words) and no opportunity to clarify with participants. Focus groups can be effective for quickly gathering input from a group of participants. However, social dynamics may result in one or a few people dominating the discussion, or “group think,” when people agree with one another rather than providing their own unique perspectives. Interviews with individuals can be a rich and varied data source because each participant has time and space to offer their own distinct perspective. Interviews also allow for follow-up questions that are difficult through survey methods. Yet, conducting interviews skillfully—avoiding leading questions and ensuring that the line of questioning yields the desired data—takes a lot of thought and practice. Kvale (1996) offers detailed guidance on how to design and carry out research interviews. Observing an expert interviewer and having them observe and give feedback as you interview can help improve your skills. Audio and video recordings of learning experiences like class sessions or group work can provide a plethora of information (e.g., verbal and nonverbal exchanges among students or between students and instructors) in a more natural setting than surveys or interviews. Yet deciding what information will serve as data to answer your research question, or how that large body of data will be systematically analyzed, can be cumbersome.
Regardless of the data collection method, you’ll need to decide how much data to collect. There is no one right sample size. A good rule of thumb is collecting data until you reach “saturation,” which is the notion that the same ideas are coming up repeatedly and that no new ideas are emerging during data collection. This means that your data collection and analysis are likely to overlap in time, with some data collection then some analysis and then more data collection.
Analytic methods in qualitative research vary widely in their interpretive complexity. As natural scientists, we favor sticking close to the data and analyzing using a method called qualitative content analysis. Content analysis involves taking quotes or segments of text and capturing their meaning with short words or phrases called codes. The process of developing codes and systematically applying them to a dataset is called coding. Coding is highly iterative and time-consuming because it typically requires multiple, careful passes through the dataset to ensure all codes have been evenly applied to all data. In a recent study, we spent 10 to 15 person-hours to code a single interview, and about 400 person-hours to complete coding for a 30-participant study. The time involved in coding depends on what is being studied, the type of coding, and who is coding the data. Saldaña (2016) provides excellent guidance on the coding process, including various ways of making sense of codes by grouping them into themes. Content analysis is just one approach to qualitative data analysis. We encourage you to learn more about different forms of qualitative approaches and choose what works best for you, including your skill level, research goals, and data (e.g., Creswell and Poth 2016; Starks and Brown Trinidad 2007).
Interpret and Write Results
There are many ways to effectively write up results, often called findings, from qualitative research. Because qualitative research involves extensive interpretation, it can sometimes be easier to integrate the results and discussion of a qualitative paper. Integration allows the interpretation (discussion) to be directly supported by the evidence in the form of quotations (results). The conclusions of the paper should avoid repeating the results and instead comment on the implications and applications of the findings: why they matter and what to do as a result. Because qualitative data are quotations rather than numbers, qualitative papers tend to be longer than papers presenting quantitative studies. That said, qualitative papers should still aim to be succinct. For instance, depending on the approach and methods, quotations can be lightly edited to remove extra words or filler language (e.g., um, uh) that is a natural part of language but otherwise irrelevant to the findings. Presenting only the most pertinent part of a quotation not only facilitates succinctness, but helps readers attend to the specific evidence that supports the claims being made. Another strategy to shorten qualitative papers is to present some findings in supplemental materials.
Final Recommendations
In closing our article, we offer some advice that we wish we knew when we began conducting qualitative research. We hope that these recommendations will help you think through issues that are likely emerge as you delve deeper into qualitative analysis, both as a producer and a consumer of qualitative research.
Consensus Coding in Qualitative Analysis
In qualitative analysis, we work to ensure that the analysis yields trustworthy findings by coding to consensus, meaning that the analytic team reaches 100 percent agreement on the application of each code to the data. Any disagreement between coders is discussed until a resolution is resolved. In some cases, these discussions may result in a code description being redefined. Redefinition of a code requires that all data previously coded using the original code be reanalyzed to ensure fit with the revised definition. As you might imagine, coding to consensus can be time-consuming. Yet, in our experience, the time invested in coding to consensus is well spent because the analysis yields deeper insights about the data and phenomenon being investigated. We also see coding to consensus as a great way to take advantage of the diverse viewpoints that team members bring to our research. By coding to consensus, we consider multiple interpretations of the data throughout the analysis process. We are well-positioned to develop theory (as appropriate for our study design) as a team because we all have engaged in meaningful conversations about our findings throughout analysis.
Some qualitative research relies on a calculated measure of intercoder reliability (ICR) instead of coding to consensus. ICR values indicate how often a set of coders agree on the application of a code in the dataset. This quantification of coding is tempting because we love numbers, yet it can also be problematic (O’Connor and Joffe 2020). For instance, aiming for high ICR can create situations when coders are pressured to agree with each other rather than bringing their own unique perspective to the coding process (e.g., Belur et al. 2018; Morse 1997). Quantifying qualitative work also can imply a false precision in the analysis. In some research, ICR is calculated partway through the analysis to determine whether an “acceptable” level of agreement has been reached, at which point the remainder of the data are coded by just one researcher. This approach of using ICR as a cut-off runs counter to what many argue is the value of qualitative research: generating new theoretical understandings informed by multiple perspectives.
Using Numbers in Qualitative Analysis
Although numbers certainly have a place in qualitative analysis (Sandelowski 2001), we encourage researchers to move beyond word clouds or frequency counts of codes and themes in their results for two reasons. First, a code or theme that is infrequently observed in the data set can still be important to the phenomenon being studied. As an analogy, consider making qualitative observations of living cells under a typical light microscope. We would most frequently see a relatively stationary cell that is punctuated by a relatively rare cell division or mitosis. If we only reported stationary observations in findings, we would overlook describing mitosis, one of the most dynamic and fundamental processes that cells display. Second, given limited sample sizes, it may be that a unique and important code or theme is reported by only one participant in the data set. In fact, rare observations can serve as “a-ha moments” that lead to a more comprehensive understanding of the phenomenon under investigation. These rare observations also may inspire new studies about topics that were not initially anticipated; this speaks to the value of qualitative research.
Closing Thoughts
We encourage readers to continue to learn about qualitative research as there is much that could not be addressed in a single article. For instance, we did not introduce how philosophical stances, like how someone views the nature of truth or what counts as evidence, influence the research process. (Creswell and Poth 2016). For now, we will close with one final piece of advice. We both became better qualitative researchers by working with mentors and collaborators who have this expertise. We encourage you to find colleagues in your networks or at your institutions who may be interested in being a collaborator, mentor, or critical friend. The complexity of students and their experiences lend themselves to qualitative approaches. We hope this article might serve as an impetus for you to learn more about qualitative research and even start your own investigations.
Data Availability Statement
The data included in this commentary have been published in an open-access journal under a Creative Commons license. Citations are included in the text.
Institutional Review Board Statement
Not applicable.
Conflict of Interest Statement
The authors have no conflicts of interest to report.
Acknowledgments
This material is based upon work supported by the National Science Foundation under award number OCE-2019589. This is the National Science Foundation’s Center for Chemical Currencies of a Microbial Planet (C-Comp) publication #026. Any opinions, findings, and conclusions or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the National Science Foundation. We thank Patricia Mabrouk for inviting us to contribute this commentary. We thank members of the Biology Education Research Group at the University of Georgia and Daniel Dries, Joseph Provost, and Verónica Segarra for their thoughtful feedback on manuscript drafts.
Agee, Jane. 2009. “Developing Qualitative Research Questions: A Reflective Process.” International Journal of Qualitative Studies in Education 22: 431–447. doi: 10.1080/09518390902736512
Aikens, Melissa L., Melissa M. Robertson, Sona Sadselia, Keiana Watkins, Mara Evans, Christopher R. Runyon, Lillian T. Eby, and Erin L. Dolan. 2017. “Race and Gender Differences in Undergraduate Research Mentoring Structures and Research Outcomes.” CBE—Life Sciences Education 16(2): ar34. doi:10.1187/cbe.16-07-0211
Aikens, Melissa L., Sona Sadselia, Keiana Watkins, Mara Evans, Lillian T. Eby, and Erin L. Dolan. 2016. “A Social Capital Perspective on the Mentoring of Undergraduate Life Science Researchers: An Empirical Study of Undergraduate–Postgraduate–Faculty Triads.” CBE—Life Sciences Education 15(2): ar16. doi: 10.1187/cbe.15-10-0208
Anfara, Vincent A., Kathleen M. Brown, and Terri L. Mangione. 2002. “Qualitative Analysis on Stage: Making the Research Process More Public.” Educational Researcher 31(7): 28–38. doi:10.3102/0013189X031007028
Belur, Jyoti, Lisa Tompson, Amy Thornton, and Miranda Simon. 2018. “Interrater Reliability in Systematic Review Methodology: Exploring Variation in Coder Decision-Making.” Sociological Methods & Research 50: 837–865. doi:10.1177/0049124118799372
Carlone, Heidi B., and Angela Johnson. 2007. “Understanding the Science Experiences of Successful Women of Color: Science Identity as an Analytic Lens.” Journal of Research in Science Teaching 44: 1187–1218. doi: 10.1002/tea.20237
Castillo-Montoya, Milagros. 2016. “Preparing for Interview Research: The Interview Protocol Refinement Framework.” Qualitative Report 21: 811–831. doi: 10.46743/2160-3715/2016.2337
Charmaz, Kathy. 2006. Constructing Grounded Theory: A Practical Guide through Qualitative Analysis . London: Sage.
Creswell, John W., and Cheryl N. Poth. 2016. Qualitative Inquiry and Research Design: Choosing among Five Approaches . Sage.
Dolan, Erin L. 2013. “Biology Education Scholarship.” IBiology. https://www.ibiology.org/career-exploration/biology-educationscholarship
Gentile, Jim, Kerry Brenner, and Amy Stephens, eds. 2017. Undergraduate Research Experiences for STEM Students: Successes, Challenges, and Opportunities . Washington, DC: National Academies Press. https://www.nap.edu/catalog/24622/undergraduate-research-experiences-for-stem-students-successes-challenges-and-opportunities
Gioia, Denny, Kevin Corley, Kathleen Eisenhardt, Martha Feldman, Ann Langley, Jane Lê, Karen Golden-Biddle, et al. 2022. “A Curated Debate: On Using ‘Templates’ in Qualitative Research.” Journal of Management Inquiry 31: 231–52. doi:10.1177/10564926221098955
Goldberg, Abbie E., and Katherine R. Allen. 2015. “Communicating Qualitative Research: Some Practical Guideposts for Scholars.” Journal of Marriage and Family 77 (1): 3–22. doi:10.1111/jomf.12153
Grant, Cynthia, and Azadeh Osanloo. 2014. “Understanding, Selecting, and Integrating a Theoretical Framework in Dissertation Research: Creating the Blueprint for Your ‘House.’” Administrative Issues Journal 4(2): 4. https://dc.swosu.edu/aij/vol4/iss2/4
Hampton, Cynthia, David Reeping, and Desen Sevi Ozkan. 2021. “Positionality Statements in Engineering Education Research: A Look at the Hand That Guides the Methodological Tools.” Studies in Engineering Education 1(2): 126–141. doi: 10.21061/see.13
Hazari, Zahra, Deepa Chari, Geoff Potvin, and Eric Brewe. 2020. “The Context Dependence of Physics Identity: Examining the Role of Performance/Competence, Recognition, Interest, and Sense of Belonging for Lower and Upper Female Physics Undergraduates.” Journal of Research in Science Teaching 57:1583–1607. doi: 10.1002/tea.21644
Hazari, Zahra, Gerhard Sonnert, Philip M. Sadler, and Marie-Claire Shanahan. 2010. “Connecting High School Physics Experiences, Outcome Expectations, Physics Identity, and Physics Career Choice: A Gender Study.” Journal of Research in Science Teaching 47: 978–1003. doi: 10.1002/tea.20363
Holmes, Andrew, and Gary Darwin. 2020. “Researcher Positionality: A Consideration of Its Influence and Place in Qualitative Research; A New Researcher Guide.” Shanlax International Journal of Education 8(4): 1–10. doi: 10.34293/education.v8i4.3232
Hunter, Anne-Barrie, Sandra L. Laursen, and Elaine Seymour. 2007. “Becoming a Scientist: The Role of Undergraduate Research in Students’ Cognitive, Personal, and Professional Development.” Science Education 91: 36–74. doi: 10.1002/sce.20173
Joshi, Megha, Melissa L. Aikens, and Erin L. Dolan. 2019. “Direct Ties to a Faculty Mentor Related to Positive Outcomes for Undergraduate Researchers.” BioScience 69: 389–397. doi10.1093/biosci/biz039
Kim, Ann Y., and Gale M. Sinatra. 2018. “Science Identity Development: An Interactionist Approach.” International Journal of STEM Education 5: 51. doi: 10.1186/s40594-018-0149-9
Knott, Eleanor, Aliya Hamid Rao, Kate Summers, and Chana Teeger. 2022. “Interviews in the Social Sciences.” Nature Reviews Methods Primers 2: 73. doi: 10.1038/s43586-022-00150-6
Korstjens, Irene, and Albine Moser. 2017. “Series: Practical Guidance to Qualitative Research. Part 2: Context, Research Questions and Designs.” European Journal of General Practice 23: 274–279. doi: 10.1080/13814788.2017.1375090
Kvale, Steinar. 1996. InterViews: An Introduction to Qualitative Research Interviewing . Thousand Oaks, CA: Sage.
Kyngäs, Helvi, Kristina Mikkonen, and Maria Kääriäinen, eds. 2020. The Application of Content Analysis in Nursing Science Research . Cham: Springer International. doi: 10.1007/978-3-030-30199-6
Limeri, Lisa B., Muhammad Zaka Asif, Benjamin H. T. Bridges, David Esparza, Trevor T. Tuma, Daquan Sanders, Alexander J. Morrison, Pallavi Rao, Joseph A. Harsh, and Adam V. Maltese. 2019. “‘Where’s My Mentor?!’ Characterizing Negative Mentoring Experiences in Undergraduate Life Science Research.” CBE—Life Sciences Education 18(4): ar61. doi: 10.1187/cbe.19-02-0036
Lincoln, Yvonna S., and Egon G. Guba. 1985. Naturalistic Inquiry . Sage.
Loeb, Susanna, Susan Dynarski, Daniel McFarland, Pamela Morris, Sean Reardon, and Sarah Reber. 2017. “Descriptive Analysis in Education: A Guide for Researchers.” NCEE 2017-4023. National Center for Education Evaluation and Regional Assistance.
Luft, Julie A., Sophia Jeong, Robert Idsardi, and Grant Gardner. 2022. “Literature Reviews, Theoretical Frameworks, and Conceptual Frameworks: An Introduction for New Biology Education Researchers.” CBE—Life Sciences Education 21(3): rm33. doi: 10.1187/cbe.21-05-0134
Majocha, Megan, Zachary Davenport, Derek C. Braun, and Cara Gormally. 2018. “‘Everyone Was Nice . . . But I Was Still Left Out’: An Interview Study about Deaf Interns’ Research Experiences in STEM.” Journal of Microbiology & Biology Education 19(1): 19.1.10. doi: 10.1128/jmbe.v19i1.1381
Mays, Nicholas, and Catherine Pope. 2020. “Quality in Qualitative Research.” In Qualitative Research in Health Care , ed. Catherine Pope and Nicholas Mays, 211–233. doi:10.1002/9781119410867.ch15
McGee, Ebony O., and Lydia Bentley. 2017. “The Troubled Success of Black Women in STEM.” Cognition and Instruction 35: 265–289. doi: 10.1080/07370008.2017.1355211
Merriam, Sharan B. 2014. Qualitative Research: A Guide to Design and Implementation . San Francisco: Wiley.
Miles, Matthew B., A. Michael Huberman, and Johnny Saldana. 2014. Qualitative Data Analysis: A Methods Sourcebook . 3rd ed. Thousand Oaks, CA: Sage.
Morse, Janice M. 1997. “‘Perfectly Healthy, but Dead’: The Myth of Inter-Rater Reliability.” Qualitative Health Research 7:445–47. doi: 10.1177/104973239700700401
Morse, Janice M., Michael Barrett, Maria Mayan, Karin Olson, and Jude Spiers. 2002. “Verification Strategies for Establishing Reliability and Validity in Qualitative Research.” International Journal of Qualitative Methods 1(2): 13–22. Doi:10.1177/160940690200100202
National Commission for the Protection of Human Subjects of Biomedical and Behavioral Research. 1978. “The Belmont Report: Ethical Principles and Guidelines for the Commission for the Protection of Human Subjects of Biomedical and Behavioral Research.” 3 vols. Bethesda, MD: National Commission for the Protection of Human Subjects of Biomedical and Behavioral Research. https://repository.library.georgetown.edu/handle/10822/779133
O’Connor, Cliodhna, and Helene Joffe. 2020. “Intercoder Reliability in Qualitative Research: Debates and Practical Guidelines.” International Journal of Qualitative Methods 19:1609406919899220. doi: 10.1177/1609406919899220
Pfeifer, Mariel A., Julio J. Cordero, and Julie Dangremond Stanton. 2023. “What I Wish My Instructor Knew: How Active Learning Influences the Classroom Experiences and Self-Advocacy of STEM Majors with ADHD and Specific Learning Disabilities.” CBE—Life Sciences Education 2(1): ar2. doi: 10.1187/cbe.21-12-0329
Pfeifer, Mariel A., C. J. Zajic, Jared M. Isaacs, Olivia A. Erickson, and Erin L. Dolan. 2023. “Beyond Performance, Competence, and Recognition: Forging a Science Researcher Identity in the Context of Research Training.” BioRxiv 2023.03.22.533783. doi: 10.1101/2023.03.22.533783
Potvin, Geoff, and Zahra Hazari. 2013. “The Development and Measurement of Identity across the Physical Sciences.” 2013 PERC Proceedings . American Association of Physics Teachers. https://www.compadre.org/Repository/document/ServeFile.cfm?ID=13182&DocID=3729
Pratt, Michael G., Kevin W. Rockmann, and Jeffrey B. Kaufmann. 2006. “Constructing Professional Identity: The Role of Work and Identity Learning Cycles in the Customization of Identity among Medical Residents.” Academy of Management Journal 49: 235–262. doi: 10.5465/AMJ.2006.20786060
Ritchie, Jane, Jane Lewis, Carol McNaughton Nicholls, and Rachel Ormston. 2013. Qualitative Research Practice: A Guide for Social Science Students and Researchers . Sage.
Roulston, Kathryn, Kathleen deMarrais, and Jamie B. Lewis. 2003. “Learning to Interview in the Social Sciences.” Qualitative Inquiry 9: 643–668. doi: 10.1177/1077800403252736
Saldaña, Johnny. 2016. The Coding Manual for Qualitative Researchers . 3rd ed. Los Angeles: Sage.
Sandelowski, Margarete. 1995. “Qualitative Analysis: What It Is and How to Begin.” Research in Nursing & Health 18: 371–375. doi: 10.1002/nur.4770180411
Sandelowski, Margarete. 1998. “Writing a Good Read: Strategies for Re-Presenting Qualitative Data.” Research in Nursing & Health 21: 375–382. doi: 10.1002/(SICI)1098-240X(199808)21:4<375::AID-NUR9>3.0.CO;2-C
Sandelowski, Margarete. 2001. “Real Qualitative Researchers Do Not Count: The Use of Numbers in Qualitative Research.” Research in Nursing & Health 24: 230–240. doi: 10.1002/nur.1025
Spangler, Denise A., and Steven R. Williams. 2019. “The Role of Theoretical Frameworks in Mathematics Education Research.” In Designing, Conducting, and Publishing Quality Research in Mathematics Education , ed. Keith R. Leatham, 3–16. Research in Mathematics Education. Cham: Springer International. doi:10.1007/978-3-030-23505-5_1
Starks, Helene, and Susan Brown Trinidad. 2007. “Choose Your Method: A Comparison of Phenomenology, Discourse Analysis, and Grounded Theory.” Qualitative Health Research 17: 1372–1380. doi: 10.1177/1049732307307031
Tracy, Sarah J. 2010. “Qualitative Quality: Eight ‘Big-Tent’ Criteria for Excellent Qualitative Research.” Qualitative Inquiry 16:837–851. doi: 10.1177/1077800410383121
University of California Museum of Paleontology. 2022. “Understanding Science: Science Flowchart.” UC Museum of Paleontology Understanding Science. https://undsci.berkeley.edu/science-flowchart
Vaccaro, Annemarie, Ezekiel W. Kimball, Ryan S. Wells, and Benjamin J. Ostiguy. “Researching students with disabilities: The importance of critical perspectives.” New directions for institutional research 2014, no. 163 (2015): 25-41. doi: 10.1002/ir.20084
Vasquez-Salgado, Yolanda, Tissyana C. Camacho, Isabel López, Gabriela Chavira, Carrie L. Saetermoe, and Crist Khachikian. 2023. “‘I Definitely Feel like a Scientist’: Exploring Science Identity Trajectories among Latinx Students in a Critical Race Theory–Informed Undergraduate Research Experience.” Infant and Child Development 32(3): e2371. doi: 10.1002/icd.2371
Watson, Rachel M., John D. Willford, and Mariel A. Pfeifer. 2018. “A Cultured Learning Environment: Implementing a Problem-and Service-Based Microbiology Capstone Course to Assess Process- and Skill-Based Learning Objectives.” Interdisciplinary Journal of Problem-Based Learning 12(1): article 8. doi:10.7771/1541-5015.1694
Welch, Catherine. 2018. “Good Qualitative Research: Opening up the Debate.” In Collaborative Research Design: Working with Business for Meaningful Findings , 401–412. Singapore: Springer. doi: 10.1007/978-981-10-5008-4
Yadav, Drishti. 2022. “Criteria for Good Qualitative Research: A Comprehensive Review.” Asia-Pacific Education Researcher 31 679–689. doi: 10.1007/s40299-021-00619-0
Yilmaz, Kaya. 2013. “Comparison of Quantitative and Qualitative Research Traditions: Epistemological, Theoretical, and Methodological Differences.” European Journal of Education 48: 311–325. doi: 10.1111/ejed.12014
Mariel A. Pfeifer
University of Georgia, [email protected]
Mariel A. Pfeifer is a postdoctoral researcher at the University of Georgia’s SPREE (Social Psychology of Research Experiences and Education) Lab. Her passion for biology education research was sparked by her experiences as an undergraduate teaching assistant, a pre-service science teacher, and a disability services coordinator. Soon Pfeifer will begin her new role as an assistant professor of biology at the University of Mississippi.
Erin L. Dolan is a professor of biochemistry and molecular biology and Georgia Athletic Association Professor of Innovative Science Education at the University of Georgia As a graduate student, Dolan volunteered in K–12 schools, which inspired her pursuit of a biology education career. She teaches introductory biology and her research group, the SPREE Lab, works to delineate features of undergraduate and graduate research that influence students’ career decisions.
More Articles in this Issue
Table of contents – fall 2023, quantifying equity in the american theater: student research at the intersection of performance and data analysis.
Dramatic literature courses in the undergraduate theater curriculum traditionally include the study of plays: their structure and themes. In a course titled Contemporary Female Playwrights at Davidson College, the learning goals go beyond script analysis and include strategies for documenting and redressing the underrepresentation of female and BIPOC playwrights in American theater.
Building Research Skills through an Undergraduate Research Project on Local Community
This research aims to build economic research skills and stimulate students’ interest in the local economy through data-based undergraduate research in entry-level economics courses. The authors developed two assignments and one student survey assessing students’ learning outcomes and implemented them in two introductory-level classes from fall 2019 to fall 2021. The survey responses confirmed that the assignments positively affected students’ primary research skills and increased students’ interest in local economic issues. The study also provides empirical evidence that undergraduate research can be carried out in both face-to-face and online classes. It confirms the positive contribution of exposing students to the research culture early in their academic journey by improving students’ skills in collecting, processing, and interpreting data on the local economy.
Fulfilling the Land-Grant Mission in Undergraduate Research in English Studies
The pandemic provided a natural experiment to test an alternative approach to teaching a traditional classroom-based research methods course; as teachers, we should not wait for such interventions but try out various strategies for effectiveness. Remote mentoring is entirely feasible for successful undergraduate research experiences. This is a particularly crucial finding in the humanities, which relies very much on discussion-based formats rather than lectures for its courses. Faculty have learned new technologies, such as Zoom, to ensure that meaningful interactions would occur.
Making Research Accessible for All through a Fully Online Cancer Genomics CURE
Course-based undergraduate research experiences (CUREs) can engage large numbers of students and provide a structured environment in which to learn valuable research skills. The ability to implement laboratory-based CUREs was hindered by the COVID-19 pandemic, generating a greater need for online options. A pilot study of an adaptation of a fully online cancer genomics CURE is described here. Students utilized freely available databases such as cBioPortal to develop novel scientific questions, generate and analyze data, collaborate with peers, and present their findings in an online environment. This format preserved the defining aspects of CUREs while promoting student ownership over their projects. Although the most common challenge was developing a hypothesis, students valued peer and instructor feedback throughout the process as well as flexible formats for communicating their research findings.
Laboratory-Based Undergraduate Research Experiences (LUREs): Evidence of Effectiveness from the Social Sciences
The impact of undergraduate research experiences (UREs) is supported by evidence from physical and life science fields, especially when student-apprentices work in traditional laboratories. Within social sciences specifically, some excellent student outcomes associated with UREs adhere to non–lab-based modalities like course-based research experiences (CUREs). Here, the authors evaluate the laboratory-based undergraduate research experiences (LUREs) as a potentially valuable approach for incorporating social science undergraduates in research. Using comparative analysis of survey data from students completing three types of social science-based UREs (n = 235), individual research experiences (IREs), CUREs, or LUREs, students perceived gains overall regardless of the type of experience, with some indication that LUREs are the most effective.
SUREbyts: Presenting Early-Year Undergraduate Students with Videos on Research Topics
Undergraduate research initiatives such as mentoring programs, conferences, and journals typically focus on the later stages of undergraduate studies. It is not unusual for a student to reach the final year of their program without developing their awareness of research within their discipline or their institution. SUREbyts is a project that provides first- and second-year undergraduate students with access to research through video recordings of professional researchers and research students discussing their own research, with each video structured around a research question with a set of possible solutions. This article presents the successes and challenges faced by the project’s initial implementation in six higher education institutions in Ireland and offers advice to institutions globally that are considering engaging their students with research in this way.
Systematic Review of Outcomes for Faculty Mentors in Undergraduate Research
Significant research has highlighted the benefits and outcomes of mentored research experiences for undergraduate students. Substantially less empirical research has examined the benefits and outcomes of these experiences for the other member of the mentoring dyad: the mentors themselves. To address this gap, a systematic review of 1,915 articles was conducted. After review, 16 articles were determined relevant. Articles were categorized based on design and theoretical framework. Further analysis revealed three categorizations: faculty mentor outcomes, barriers to mentorship of undergraduate research students, and supporting factors associated with mentorship of students. Results indicate that faculty mentors in undergraduate research contexts face more barriers to mentorship than supporting factors. Three lines of inquiry are proposed for future researchers.
Undergraduate Research: Why and How Do We Mentor?
Our advocacy for undergraduate research, mentorship, and equity is strengthened by being explicit about why they are vital. Research mentors guide undergraduates in learning how to learn, in evaluating evidence, and in discovering talents and career interests. Our mission can be extended by bringing a research perspective to the courses we teach. Effective mentorship involves appreciating individual differences while having shared goals. As mentors, we aim to provide undergraduates with opportunities to participate in research that can lead to discovering new knowledge. Students can build on this experience to become proactive in making a difference. The global future depends on today’s undergraduates, who are tomorrow’s decision-makers, innovators, and leaders.
Step Up for SPUR
This editorial details observations from a departing associate editor of SPUR about the experience of working on the journal. The author contextualizes this work within the academic journal editorial process, focusing on specific challenges in recruiting reviewers for the peer review process. The author stresses the importance of broad participation from the undergraduate research community, including submitting articles to SPUR , accepting requests to review manuscripts, and participating in the editorial board, all to ensure the sustainability and intellectual vitality of the journal.

SUBSCRIPTION
SPUR advances knowledge and understanding of novel and effective approaches to mentored undergraduate research, scholarship, and creative inquiry by publishing high-quality, rigorously peer reviewed studies written by scholars and practitioners of undergraduate research, scholarship, and creative inquiry. The SPUR Journal is a leading CUR member benefit. Gain access to all electronic articles by joining CUR.

Biological Literature: Quantitative vs Qualitative Research
- The Scientific Method Explained
- Parts of a Scientific Article
- How to Read Scientific Articles
- Research vs Review Articles
- Quantitative vs Qualitative Research
- How do I find a Quantitative article?
- Find Books/eBooks
- Linking Google Scholar to Gee Library Resources
- Related Websites
- Evaluating Resources
- Documenting Sources (Citations)
- Avoiding Plagiarism
- Choosing Your Topic This link opens in a new window
- Creating a Literature Review
- Related Guides
- Library Session Materials
Differences in a Nutshell
In the world of research, there are two general approaches to gathering and reporting information: qualitative and quantitative approaches. Qualitative research generates non-numerical data while quantitative research generates numerical data or information that can be converted into numbers.
Comparison of the Characteristics of Qualitative & Quantitative Articles

Page Attribution
UTA Libraries. Biological Literature: Q uantitative and Qualitative Research . October 3, 2022, 2:28 p.m. CST. URL: https://libguides.uta.edu/quantitative_and_qualitative_research/differences
How to determine which type of article you may have found
1. Does it have Method and Results sections?
EXAMPLE ABSTRACT
a) If not, the article is secondary research (typically a review) .
b) If it does have Method and Results sections, continue to step 2.
2. In the Method section , does it talk about a literature search strategy?
a) If you answered yes, i t is a systematic review or meta-analysis .
b) If you answered no, and that section describes a research study (research participants, an intervention, etc.), it is original research . Continue to step 3 to determine its type.
STEP 3: Now, let's determine what type of original research this is.
3a. In the Method section, is the study described as an interview, observation, or questionnaire? Does the study involve looking into self-reported beliefs, thoughts, etc. ?
a ) If you answered yes, it’s a qualitative study .
b) If you answered no, see 3b.
3b. In the Method section, does the study describe the use of already-existing data (for example, reviewing patient admissions from the past 3 months)?
a ) If you answered yes, it’s a retrospective study .
b) If you answered no, see 3c.
3c. In the Method or Results sections/areas , does it mention using quantitative analyses or statistical tests ( e.g. , ANOVA, t-test, p values)?
EXAMPLE ARTICLE EXCERPT
a ) If you answered yes, it’s a quantitative study .
b) If you answered no, it’s qualitative.
BONUS. Are 3a AND 3c both true? In other words, does the study use BOTH quantitative and qualitative methods?
If so, it's a mixed methods study.
- << Previous: Research vs Review Articles
- Next: How do I find a Quantitative article? >>
- Last Updated: Oct 3, 2022 2:30 PM
- URL: https://libguides.mssu.edu/c.php?g=865170
This site is maintained by the librarians of George A. Spiva Library . If you have a question or comment about the Library's LibGuides, please contact the site administrator .
- Open access
- Published: 27 May 2020
How to use and assess qualitative research methods
- Loraine Busetto ORCID: orcid.org/0000-0002-9228-7875 1 ,
- Wolfgang Wick 1 , 2 &
- Christoph Gumbinger 1
Neurological Research and Practice volume 2 , Article number: 14 ( 2020 ) Cite this article
765k Accesses
345 Citations
90 Altmetric
Metrics details
This paper aims to provide an overview of the use and assessment of qualitative research methods in the health sciences. Qualitative research can be defined as the study of the nature of phenomena and is especially appropriate for answering questions of why something is (not) observed, assessing complex multi-component interventions, and focussing on intervention improvement. The most common methods of data collection are document study, (non-) participant observations, semi-structured interviews and focus groups. For data analysis, field-notes and audio-recordings are transcribed into protocols and transcripts, and coded using qualitative data management software. Criteria such as checklists, reflexivity, sampling strategies, piloting, co-coding, member-checking and stakeholder involvement can be used to enhance and assess the quality of the research conducted. Using qualitative in addition to quantitative designs will equip us with better tools to address a greater range of research problems, and to fill in blind spots in current neurological research and practice.
The aim of this paper is to provide an overview of qualitative research methods, including hands-on information on how they can be used, reported and assessed. This article is intended for beginning qualitative researchers in the health sciences as well as experienced quantitative researchers who wish to broaden their understanding of qualitative research.
What is qualitative research?
Qualitative research is defined as “the study of the nature of phenomena”, including “their quality, different manifestations, the context in which they appear or the perspectives from which they can be perceived” , but excluding “their range, frequency and place in an objectively determined chain of cause and effect” [ 1 ]. This formal definition can be complemented with a more pragmatic rule of thumb: qualitative research generally includes data in form of words rather than numbers [ 2 ].
Why conduct qualitative research?
Because some research questions cannot be answered using (only) quantitative methods. For example, one Australian study addressed the issue of why patients from Aboriginal communities often present late or not at all to specialist services offered by tertiary care hospitals. Using qualitative interviews with patients and staff, it found one of the most significant access barriers to be transportation problems, including some towns and communities simply not having a bus service to the hospital [ 3 ]. A quantitative study could have measured the number of patients over time or even looked at possible explanatory factors – but only those previously known or suspected to be of relevance. To discover reasons for observed patterns, especially the invisible or surprising ones, qualitative designs are needed.
While qualitative research is common in other fields, it is still relatively underrepresented in health services research. The latter field is more traditionally rooted in the evidence-based-medicine paradigm, as seen in " research that involves testing the effectiveness of various strategies to achieve changes in clinical practice, preferably applying randomised controlled trial study designs (...) " [ 4 ]. This focus on quantitative research and specifically randomised controlled trials (RCT) is visible in the idea of a hierarchy of research evidence which assumes that some research designs are objectively better than others, and that choosing a "lesser" design is only acceptable when the better ones are not practically or ethically feasible [ 5 , 6 ]. Others, however, argue that an objective hierarchy does not exist, and that, instead, the research design and methods should be chosen to fit the specific research question at hand – "questions before methods" [ 2 , 7 , 8 , 9 ]. This means that even when an RCT is possible, some research problems require a different design that is better suited to addressing them. Arguing in JAMA, Berwick uses the example of rapid response teams in hospitals, which he describes as " a complex, multicomponent intervention – essentially a process of social change" susceptible to a range of different context factors including leadership or organisation history. According to him, "[in] such complex terrain, the RCT is an impoverished way to learn. Critics who use it as a truth standard in this context are incorrect" [ 8 ] . Instead of limiting oneself to RCTs, Berwick recommends embracing a wider range of methods , including qualitative ones, which for "these specific applications, (...) are not compromises in learning how to improve; they are superior" [ 8 ].
Research problems that can be approached particularly well using qualitative methods include assessing complex multi-component interventions or systems (of change), addressing questions beyond “what works”, towards “what works for whom when, how and why”, and focussing on intervention improvement rather than accreditation [ 7 , 9 , 10 , 11 , 12 ]. Using qualitative methods can also help shed light on the “softer” side of medical treatment. For example, while quantitative trials can measure the costs and benefits of neuro-oncological treatment in terms of survival rates or adverse effects, qualitative research can help provide a better understanding of patient or caregiver stress, visibility of illness or out-of-pocket expenses.
How to conduct qualitative research?
Given that qualitative research is characterised by flexibility, openness and responsivity to context, the steps of data collection and analysis are not as separate and consecutive as they tend to be in quantitative research [ 13 , 14 ]. As Fossey puts it : “sampling, data collection, analysis and interpretation are related to each other in a cyclical (iterative) manner, rather than following one after another in a stepwise approach” [ 15 ]. The researcher can make educated decisions with regard to the choice of method, how they are implemented, and to which and how many units they are applied [ 13 ]. As shown in Fig. 1 , this can involve several back-and-forth steps between data collection and analysis where new insights and experiences can lead to adaption and expansion of the original plan. Some insights may also necessitate a revision of the research question and/or the research design as a whole. The process ends when saturation is achieved, i.e. when no relevant new information can be found (see also below: sampling and saturation). For reasons of transparency, it is essential for all decisions as well as the underlying reasoning to be well-documented.

Iterative research process
While it is not always explicitly addressed, qualitative methods reflect a different underlying research paradigm than quantitative research (e.g. constructivism or interpretivism as opposed to positivism). The choice of methods can be based on the respective underlying substantive theory or theoretical framework used by the researcher [ 2 ].
Data collection
The methods of qualitative data collection most commonly used in health research are document study, observations, semi-structured interviews and focus groups [ 1 , 14 , 16 , 17 ].
Document study
Document study (also called document analysis) refers to the review by the researcher of written materials [ 14 ]. These can include personal and non-personal documents such as archives, annual reports, guidelines, policy documents, diaries or letters.
Observations
Observations are particularly useful to gain insights into a certain setting and actual behaviour – as opposed to reported behaviour or opinions [ 13 ]. Qualitative observations can be either participant or non-participant in nature. In participant observations, the observer is part of the observed setting, for example a nurse working in an intensive care unit [ 18 ]. In non-participant observations, the observer is “on the outside looking in”, i.e. present in but not part of the situation, trying not to influence the setting by their presence. Observations can be planned (e.g. for 3 h during the day or night shift) or ad hoc (e.g. as soon as a stroke patient arrives at the emergency room). During the observation, the observer takes notes on everything or certain pre-determined parts of what is happening around them, for example focusing on physician-patient interactions or communication between different professional groups. Written notes can be taken during or after the observations, depending on feasibility (which is usually lower during participant observations) and acceptability (e.g. when the observer is perceived to be judging the observed). Afterwards, these field notes are transcribed into observation protocols. If more than one observer was involved, field notes are taken independently, but notes can be consolidated into one protocol after discussions. Advantages of conducting observations include minimising the distance between the researcher and the researched, the potential discovery of topics that the researcher did not realise were relevant and gaining deeper insights into the real-world dimensions of the research problem at hand [ 18 ].
Semi-structured interviews
Hijmans & Kuyper describe qualitative interviews as “an exchange with an informal character, a conversation with a goal” [ 19 ]. Interviews are used to gain insights into a person’s subjective experiences, opinions and motivations – as opposed to facts or behaviours [ 13 ]. Interviews can be distinguished by the degree to which they are structured (i.e. a questionnaire), open (e.g. free conversation or autobiographical interviews) or semi-structured [ 2 , 13 ]. Semi-structured interviews are characterized by open-ended questions and the use of an interview guide (or topic guide/list) in which the broad areas of interest, sometimes including sub-questions, are defined [ 19 ]. The pre-defined topics in the interview guide can be derived from the literature, previous research or a preliminary method of data collection, e.g. document study or observations. The topic list is usually adapted and improved at the start of the data collection process as the interviewer learns more about the field [ 20 ]. Across interviews the focus on the different (blocks of) questions may differ and some questions may be skipped altogether (e.g. if the interviewee is not able or willing to answer the questions or for concerns about the total length of the interview) [ 20 ]. Qualitative interviews are usually not conducted in written format as it impedes on the interactive component of the method [ 20 ]. In comparison to written surveys, qualitative interviews have the advantage of being interactive and allowing for unexpected topics to emerge and to be taken up by the researcher. This can also help overcome a provider or researcher-centred bias often found in written surveys, which by nature, can only measure what is already known or expected to be of relevance to the researcher. Interviews can be audio- or video-taped; but sometimes it is only feasible or acceptable for the interviewer to take written notes [ 14 , 16 , 20 ].
Focus groups
Focus groups are group interviews to explore participants’ expertise and experiences, including explorations of how and why people behave in certain ways [ 1 ]. Focus groups usually consist of 6–8 people and are led by an experienced moderator following a topic guide or “script” [ 21 ]. They can involve an observer who takes note of the non-verbal aspects of the situation, possibly using an observation guide [ 21 ]. Depending on researchers’ and participants’ preferences, the discussions can be audio- or video-taped and transcribed afterwards [ 21 ]. Focus groups are useful for bringing together homogeneous (to a lesser extent heterogeneous) groups of participants with relevant expertise and experience on a given topic on which they can share detailed information [ 21 ]. Focus groups are a relatively easy, fast and inexpensive method to gain access to information on interactions in a given group, i.e. “the sharing and comparing” among participants [ 21 ]. Disadvantages include less control over the process and a lesser extent to which each individual may participate. Moreover, focus group moderators need experience, as do those tasked with the analysis of the resulting data. Focus groups can be less appropriate for discussing sensitive topics that participants might be reluctant to disclose in a group setting [ 13 ]. Moreover, attention must be paid to the emergence of “groupthink” as well as possible power dynamics within the group, e.g. when patients are awed or intimidated by health professionals.
Choosing the “right” method
As explained above, the school of thought underlying qualitative research assumes no objective hierarchy of evidence and methods. This means that each choice of single or combined methods has to be based on the research question that needs to be answered and a critical assessment with regard to whether or to what extent the chosen method can accomplish this – i.e. the “fit” between question and method [ 14 ]. It is necessary for these decisions to be documented when they are being made, and to be critically discussed when reporting methods and results.
Let us assume that our research aim is to examine the (clinical) processes around acute endovascular treatment (EVT), from the patient’s arrival at the emergency room to recanalization, with the aim to identify possible causes for delay and/or other causes for sub-optimal treatment outcome. As a first step, we could conduct a document study of the relevant standard operating procedures (SOPs) for this phase of care – are they up-to-date and in line with current guidelines? Do they contain any mistakes, irregularities or uncertainties that could cause delays or other problems? Regardless of the answers to these questions, the results have to be interpreted based on what they are: a written outline of what care processes in this hospital should look like. If we want to know what they actually look like in practice, we can conduct observations of the processes described in the SOPs. These results can (and should) be analysed in themselves, but also in comparison to the results of the document analysis, especially as regards relevant discrepancies. Do the SOPs outline specific tests for which no equipment can be observed or tasks to be performed by specialized nurses who are not present during the observation? It might also be possible that the written SOP is outdated, but the actual care provided is in line with current best practice. In order to find out why these discrepancies exist, it can be useful to conduct interviews. Are the physicians simply not aware of the SOPs (because their existence is limited to the hospital’s intranet) or do they actively disagree with them or does the infrastructure make it impossible to provide the care as described? Another rationale for adding interviews is that some situations (or all of their possible variations for different patient groups or the day, night or weekend shift) cannot practically or ethically be observed. In this case, it is possible to ask those involved to report on their actions – being aware that this is not the same as the actual observation. A senior physician’s or hospital manager’s description of certain situations might differ from a nurse’s or junior physician’s one, maybe because they intentionally misrepresent facts or maybe because different aspects of the process are visible or important to them. In some cases, it can also be relevant to consider to whom the interviewee is disclosing this information – someone they trust, someone they are otherwise not connected to, or someone they suspect or are aware of being in a potentially “dangerous” power relationship to them. Lastly, a focus group could be conducted with representatives of the relevant professional groups to explore how and why exactly they provide care around EVT. The discussion might reveal discrepancies (between SOPs and actual care or between different physicians) and motivations to the researchers as well as to the focus group members that they might not have been aware of themselves. For the focus group to deliver relevant information, attention has to be paid to its composition and conduct, for example, to make sure that all participants feel safe to disclose sensitive or potentially problematic information or that the discussion is not dominated by (senior) physicians only. The resulting combination of data collection methods is shown in Fig. 2 .

Possible combination of data collection methods
Attributions for icons: “Book” by Serhii Smirnov, “Interview” by Adrien Coquet, FR, “Magnifying Glass” by anggun, ID, “Business communication” by Vectors Market; all from the Noun Project
The combination of multiple data source as described for this example can be referred to as “triangulation”, in which multiple measurements are carried out from different angles to achieve a more comprehensive understanding of the phenomenon under study [ 22 , 23 ].
Data analysis
To analyse the data collected through observations, interviews and focus groups these need to be transcribed into protocols and transcripts (see Fig. 3 ). Interviews and focus groups can be transcribed verbatim , with or without annotations for behaviour (e.g. laughing, crying, pausing) and with or without phonetic transcription of dialects and filler words, depending on what is expected or known to be relevant for the analysis. In the next step, the protocols and transcripts are coded , that is, marked (or tagged, labelled) with one or more short descriptors of the content of a sentence or paragraph [ 2 , 15 , 23 ]. Jansen describes coding as “connecting the raw data with “theoretical” terms” [ 20 ]. In a more practical sense, coding makes raw data sortable. This makes it possible to extract and examine all segments describing, say, a tele-neurology consultation from multiple data sources (e.g. SOPs, emergency room observations, staff and patient interview). In a process of synthesis and abstraction, the codes are then grouped, summarised and/or categorised [ 15 , 20 ]. The end product of the coding or analysis process is a descriptive theory of the behavioural pattern under investigation [ 20 ]. The coding process is performed using qualitative data management software, the most common ones being InVivo, MaxQDA and Atlas.ti. It should be noted that these are data management tools which support the analysis performed by the researcher(s) [ 14 ].

From data collection to data analysis
Attributions for icons: see Fig. 2 , also “Speech to text” by Trevor Dsouza, “Field Notes” by Mike O’Brien, US, “Voice Record” by ProSymbols, US, “Inspection” by Made, AU, and “Cloud” by Graphic Tigers; all from the Noun Project
How to report qualitative research?
Protocols of qualitative research can be published separately and in advance of the study results. However, the aim is not the same as in RCT protocols, i.e. to pre-define and set in stone the research questions and primary or secondary endpoints. Rather, it is a way to describe the research methods in detail, which might not be possible in the results paper given journals’ word limits. Qualitative research papers are usually longer than their quantitative counterparts to allow for deep understanding and so-called “thick description”. In the methods section, the focus is on transparency of the methods used, including why, how and by whom they were implemented in the specific study setting, so as to enable a discussion of whether and how this may have influenced data collection, analysis and interpretation. The results section usually starts with a paragraph outlining the main findings, followed by more detailed descriptions of, for example, the commonalities, discrepancies or exceptions per category [ 20 ]. Here it is important to support main findings by relevant quotations, which may add information, context, emphasis or real-life examples [ 20 , 23 ]. It is subject to debate in the field whether it is relevant to state the exact number or percentage of respondents supporting a certain statement (e.g. “Five interviewees expressed negative feelings towards XYZ”) [ 21 ].
How to combine qualitative with quantitative research?
Qualitative methods can be combined with other methods in multi- or mixed methods designs, which “[employ] two or more different methods [ …] within the same study or research program rather than confining the research to one single method” [ 24 ]. Reasons for combining methods can be diverse, including triangulation for corroboration of findings, complementarity for illustration and clarification of results, expansion to extend the breadth and range of the study, explanation of (unexpected) results generated with one method with the help of another, or offsetting the weakness of one method with the strength of another [ 1 , 17 , 24 , 25 , 26 ]. The resulting designs can be classified according to when, why and how the different quantitative and/or qualitative data strands are combined. The three most common types of mixed method designs are the convergent parallel design , the explanatory sequential design and the exploratory sequential design. The designs with examples are shown in Fig. 4 .

Three common mixed methods designs
In the convergent parallel design, a qualitative study is conducted in parallel to and independently of a quantitative study, and the results of both studies are compared and combined at the stage of interpretation of results. Using the above example of EVT provision, this could entail setting up a quantitative EVT registry to measure process times and patient outcomes in parallel to conducting the qualitative research outlined above, and then comparing results. Amongst other things, this would make it possible to assess whether interview respondents’ subjective impressions of patients receiving good care match modified Rankin Scores at follow-up, or whether observed delays in care provision are exceptions or the rule when compared to door-to-needle times as documented in the registry. In the explanatory sequential design, a quantitative study is carried out first, followed by a qualitative study to help explain the results from the quantitative study. This would be an appropriate design if the registry alone had revealed relevant delays in door-to-needle times and the qualitative study would be used to understand where and why these occurred, and how they could be improved. In the exploratory design, the qualitative study is carried out first and its results help informing and building the quantitative study in the next step [ 26 ]. If the qualitative study around EVT provision had shown a high level of dissatisfaction among the staff members involved, a quantitative questionnaire investigating staff satisfaction could be set up in the next step, informed by the qualitative study on which topics dissatisfaction had been expressed. Amongst other things, the questionnaire design would make it possible to widen the reach of the research to more respondents from different (types of) hospitals, regions, countries or settings, and to conduct sub-group analyses for different professional groups.
How to assess qualitative research?
A variety of assessment criteria and lists have been developed for qualitative research, ranging in their focus and comprehensiveness [ 14 , 17 , 27 ]. However, none of these has been elevated to the “gold standard” in the field. In the following, we therefore focus on a set of commonly used assessment criteria that, from a practical standpoint, a researcher can look for when assessing a qualitative research report or paper.
Assessors should check the authors’ use of and adherence to the relevant reporting checklists (e.g. Standards for Reporting Qualitative Research (SRQR)) to make sure all items that are relevant for this type of research are addressed [ 23 , 28 ]. Discussions of quantitative measures in addition to or instead of these qualitative measures can be a sign of lower quality of the research (paper). Providing and adhering to a checklist for qualitative research contributes to an important quality criterion for qualitative research, namely transparency [ 15 , 17 , 23 ].
Reflexivity
While methodological transparency and complete reporting is relevant for all types of research, some additional criteria must be taken into account for qualitative research. This includes what is called reflexivity, i.e. sensitivity to the relationship between the researcher and the researched, including how contact was established and maintained, or the background and experience of the researcher(s) involved in data collection and analysis. Depending on the research question and population to be researched this can be limited to professional experience, but it may also include gender, age or ethnicity [ 17 , 27 ]. These details are relevant because in qualitative research, as opposed to quantitative research, the researcher as a person cannot be isolated from the research process [ 23 ]. It may influence the conversation when an interviewed patient speaks to an interviewer who is a physician, or when an interviewee is asked to discuss a gynaecological procedure with a male interviewer, and therefore the reader must be made aware of these details [ 19 ].
Sampling and saturation
The aim of qualitative sampling is for all variants of the objects of observation that are deemed relevant for the study to be present in the sample “ to see the issue and its meanings from as many angles as possible” [ 1 , 16 , 19 , 20 , 27 ] , and to ensure “information-richness [ 15 ]. An iterative sampling approach is advised, in which data collection (e.g. five interviews) is followed by data analysis, followed by more data collection to find variants that are lacking in the current sample. This process continues until no new (relevant) information can be found and further sampling becomes redundant – which is called saturation [ 1 , 15 ] . In other words: qualitative data collection finds its end point not a priori , but when the research team determines that saturation has been reached [ 29 , 30 ].
This is also the reason why most qualitative studies use deliberate instead of random sampling strategies. This is generally referred to as “ purposive sampling” , in which researchers pre-define which types of participants or cases they need to include so as to cover all variations that are expected to be of relevance, based on the literature, previous experience or theory (i.e. theoretical sampling) [ 14 , 20 ]. Other types of purposive sampling include (but are not limited to) maximum variation sampling, critical case sampling or extreme or deviant case sampling [ 2 ]. In the above EVT example, a purposive sample could include all relevant professional groups and/or all relevant stakeholders (patients, relatives) and/or all relevant times of observation (day, night and weekend shift).
Assessors of qualitative research should check whether the considerations underlying the sampling strategy were sound and whether or how researchers tried to adapt and improve their strategies in stepwise or cyclical approaches between data collection and analysis to achieve saturation [ 14 ].
Good qualitative research is iterative in nature, i.e. it goes back and forth between data collection and analysis, revising and improving the approach where necessary. One example of this are pilot interviews, where different aspects of the interview (especially the interview guide, but also, for example, the site of the interview or whether the interview can be audio-recorded) are tested with a small number of respondents, evaluated and revised [ 19 ]. In doing so, the interviewer learns which wording or types of questions work best, or which is the best length of an interview with patients who have trouble concentrating for an extended time. Of course, the same reasoning applies to observations or focus groups which can also be piloted.
Ideally, coding should be performed by at least two researchers, especially at the beginning of the coding process when a common approach must be defined, including the establishment of a useful coding list (or tree), and when a common meaning of individual codes must be established [ 23 ]. An initial sub-set or all transcripts can be coded independently by the coders and then compared and consolidated after regular discussions in the research team. This is to make sure that codes are applied consistently to the research data.
Member checking
Member checking, also called respondent validation , refers to the practice of checking back with study respondents to see if the research is in line with their views [ 14 , 27 ]. This can happen after data collection or analysis or when first results are available [ 23 ]. For example, interviewees can be provided with (summaries of) their transcripts and asked whether they believe this to be a complete representation of their views or whether they would like to clarify or elaborate on their responses [ 17 ]. Respondents’ feedback on these issues then becomes part of the data collection and analysis [ 27 ].
Stakeholder involvement
In those niches where qualitative approaches have been able to evolve and grow, a new trend has seen the inclusion of patients and their representatives not only as study participants (i.e. “members”, see above) but as consultants to and active participants in the broader research process [ 31 , 32 , 33 ]. The underlying assumption is that patients and other stakeholders hold unique perspectives and experiences that add value beyond their own single story, making the research more relevant and beneficial to researchers, study participants and (future) patients alike [ 34 , 35 ]. Using the example of patients on or nearing dialysis, a recent scoping review found that 80% of clinical research did not address the top 10 research priorities identified by patients and caregivers [ 32 , 36 ]. In this sense, the involvement of the relevant stakeholders, especially patients and relatives, is increasingly being seen as a quality indicator in and of itself.
How not to assess qualitative research
The above overview does not include certain items that are routine in assessments of quantitative research. What follows is a non-exhaustive, non-representative, experience-based list of the quantitative criteria often applied to the assessment of qualitative research, as well as an explanation of the limited usefulness of these endeavours.
Protocol adherence
Given the openness and flexibility of qualitative research, it should not be assessed by how well it adheres to pre-determined and fixed strategies – in other words: its rigidity. Instead, the assessor should look for signs of adaptation and refinement based on lessons learned from earlier steps in the research process.
Sample size
For the reasons explained above, qualitative research does not require specific sample sizes, nor does it require that the sample size be determined a priori [ 1 , 14 , 27 , 37 , 38 , 39 ]. Sample size can only be a useful quality indicator when related to the research purpose, the chosen methodology and the composition of the sample, i.e. who was included and why.
Randomisation
While some authors argue that randomisation can be used in qualitative research, this is not commonly the case, as neither its feasibility nor its necessity or usefulness has been convincingly established for qualitative research [ 13 , 27 ]. Relevant disadvantages include the negative impact of a too large sample size as well as the possibility (or probability) of selecting “ quiet, uncooperative or inarticulate individuals ” [ 17 ]. Qualitative studies do not use control groups, either.
Interrater reliability, variability and other “objectivity checks”
The concept of “interrater reliability” is sometimes used in qualitative research to assess to which extent the coding approach overlaps between the two co-coders. However, it is not clear what this measure tells us about the quality of the analysis [ 23 ]. This means that these scores can be included in qualitative research reports, preferably with some additional information on what the score means for the analysis, but it is not a requirement. Relatedly, it is not relevant for the quality or “objectivity” of qualitative research to separate those who recruited the study participants and collected and analysed the data. Experiences even show that it might be better to have the same person or team perform all of these tasks [ 20 ]. First, when researchers introduce themselves during recruitment this can enhance trust when the interview takes place days or weeks later with the same researcher. Second, when the audio-recording is transcribed for analysis, the researcher conducting the interviews will usually remember the interviewee and the specific interview situation during data analysis. This might be helpful in providing additional context information for interpretation of data, e.g. on whether something might have been meant as a joke [ 18 ].
Not being quantitative research
Being qualitative research instead of quantitative research should not be used as an assessment criterion if it is used irrespectively of the research problem at hand. Similarly, qualitative research should not be required to be combined with quantitative research per se – unless mixed methods research is judged as inherently better than single-method research. In this case, the same criterion should be applied for quantitative studies without a qualitative component.
The main take-away points of this paper are summarised in Table 1 . We aimed to show that, if conducted well, qualitative research can answer specific research questions that cannot to be adequately answered using (only) quantitative designs. Seeing qualitative and quantitative methods as equal will help us become more aware and critical of the “fit” between the research problem and our chosen methods: I can conduct an RCT to determine the reasons for transportation delays of acute stroke patients – but should I? It also provides us with a greater range of tools to tackle a greater range of research problems more appropriately and successfully, filling in the blind spots on one half of the methodological spectrum to better address the whole complexity of neurological research and practice.
Availability of data and materials
Not applicable.
Abbreviations
Endovascular treatment
Randomised Controlled Trial
Standard Operating Procedure
Standards for Reporting Qualitative Research
Philipsen, H., & Vernooij-Dassen, M. (2007). Kwalitatief onderzoek: nuttig, onmisbaar en uitdagend. In L. PLBJ & H. TCo (Eds.), Kwalitatief onderzoek: Praktische methoden voor de medische praktijk . [Qualitative research: useful, indispensable and challenging. In: Qualitative research: Practical methods for medical practice (pp. 5–12). Houten: Bohn Stafleu van Loghum.
Chapter Google Scholar
Punch, K. F. (2013). Introduction to social research: Quantitative and qualitative approaches . London: Sage.
Kelly, J., Dwyer, J., Willis, E., & Pekarsky, B. (2014). Travelling to the city for hospital care: Access factors in country aboriginal patient journeys. Australian Journal of Rural Health, 22 (3), 109–113.
Article Google Scholar
Nilsen, P., Ståhl, C., Roback, K., & Cairney, P. (2013). Never the twain shall meet? - a comparison of implementation science and policy implementation research. Implementation Science, 8 (1), 1–12.
Howick J, Chalmers I, Glasziou, P., Greenhalgh, T., Heneghan, C., Liberati, A., Moschetti, I., Phillips, B., & Thornton, H. (2011). The 2011 Oxford CEBM evidence levels of evidence (introductory document) . Oxford Center for Evidence Based Medicine. https://www.cebm.net/2011/06/2011-oxford-cebm-levels-evidence-introductory-document/ .
Eakin, J. M. (2016). Educating critical qualitative health researchers in the land of the randomized controlled trial. Qualitative Inquiry, 22 (2), 107–118.
May, A., & Mathijssen, J. (2015). Alternatieven voor RCT bij de evaluatie van effectiviteit van interventies!? Eindrapportage. In Alternatives for RCTs in the evaluation of effectiveness of interventions!? Final report .
Google Scholar
Berwick, D. M. (2008). The science of improvement. Journal of the American Medical Association, 299 (10), 1182–1184.
Article CAS Google Scholar
Christ, T. W. (2014). Scientific-based research and randomized controlled trials, the “gold” standard? Alternative paradigms and mixed methodologies. Qualitative Inquiry, 20 (1), 72–80.
Lamont, T., Barber, N., Jd, P., Fulop, N., Garfield-Birkbeck, S., Lilford, R., Mear, L., Raine, R., & Fitzpatrick, R. (2016). New approaches to evaluating complex health and care systems. BMJ, 352:i154.
Drabble, S. J., & O’Cathain, A. (2015). Moving from Randomized Controlled Trials to Mixed Methods Intervention Evaluation. In S. Hesse-Biber & R. B. Johnson (Eds.), The Oxford Handbook of Multimethod and Mixed Methods Research Inquiry (pp. 406–425). London: Oxford University Press.
Chambers, D. A., Glasgow, R. E., & Stange, K. C. (2013). The dynamic sustainability framework: Addressing the paradox of sustainment amid ongoing change. Implementation Science : IS, 8 , 117.
Hak, T. (2007). Waarnemingsmethoden in kwalitatief onderzoek. In L. PLBJ & H. TCo (Eds.), Kwalitatief onderzoek: Praktische methoden voor de medische praktijk . [Observation methods in qualitative research] (pp. 13–25). Houten: Bohn Stafleu van Loghum.
Russell, C. K., & Gregory, D. M. (2003). Evaluation of qualitative research studies. Evidence Based Nursing, 6 (2), 36–40.
Fossey, E., Harvey, C., McDermott, F., & Davidson, L. (2002). Understanding and evaluating qualitative research. Australian and New Zealand Journal of Psychiatry, 36 , 717–732.
Yanow, D. (2000). Conducting interpretive policy analysis (Vol. 47). Thousand Oaks: Sage University Papers Series on Qualitative Research Methods.
Shenton, A. K. (2004). Strategies for ensuring trustworthiness in qualitative research projects. Education for Information, 22 , 63–75.
van der Geest, S. (2006). Participeren in ziekte en zorg: meer over kwalitatief onderzoek. Huisarts en Wetenschap, 49 (4), 283–287.
Hijmans, E., & Kuyper, M. (2007). Het halfopen interview als onderzoeksmethode. In L. PLBJ & H. TCo (Eds.), Kwalitatief onderzoek: Praktische methoden voor de medische praktijk . [The half-open interview as research method (pp. 43–51). Houten: Bohn Stafleu van Loghum.
Jansen, H. (2007). Systematiek en toepassing van de kwalitatieve survey. In L. PLBJ & H. TCo (Eds.), Kwalitatief onderzoek: Praktische methoden voor de medische praktijk . [Systematics and implementation of the qualitative survey (pp. 27–41). Houten: Bohn Stafleu van Loghum.
Pv, R., & Peremans, L. (2007). Exploreren met focusgroepgesprekken: de ‘stem’ van de groep onder de loep. In L. PLBJ & H. TCo (Eds.), Kwalitatief onderzoek: Praktische methoden voor de medische praktijk . [Exploring with focus group conversations: the “voice” of the group under the magnifying glass (pp. 53–64). Houten: Bohn Stafleu van Loghum.
Carter, N., Bryant-Lukosius, D., DiCenso, A., Blythe, J., & Neville, A. J. (2014). The use of triangulation in qualitative research. Oncology Nursing Forum, 41 (5), 545–547.
Boeije H: Analyseren in kwalitatief onderzoek: Denken en doen, [Analysis in qualitative research: Thinking and doing] vol. Den Haag Boom Lemma uitgevers; 2012.
Hunter, A., & Brewer, J. (2015). Designing Multimethod Research. In S. Hesse-Biber & R. B. Johnson (Eds.), The Oxford Handbook of Multimethod and Mixed Methods Research Inquiry (pp. 185–205). London: Oxford University Press.
Archibald, M. M., Radil, A. I., Zhang, X., & Hanson, W. E. (2015). Current mixed methods practices in qualitative research: A content analysis of leading journals. International Journal of Qualitative Methods, 14 (2), 5–33.
Creswell, J. W., & Plano Clark, V. L. (2011). Choosing a Mixed Methods Design. In Designing and Conducting Mixed Methods Research . Thousand Oaks: SAGE Publications.
Mays, N., & Pope, C. (2000). Assessing quality in qualitative research. BMJ, 320 (7226), 50–52.
O'Brien, B. C., Harris, I. B., Beckman, T. J., Reed, D. A., & Cook, D. A. (2014). Standards for reporting qualitative research: A synthesis of recommendations. Academic Medicine : Journal of the Association of American Medical Colleges, 89 (9), 1245–1251.
Saunders, B., Sim, J., Kingstone, T., Baker, S., Waterfield, J., Bartlam, B., Burroughs, H., & Jinks, C. (2018). Saturation in qualitative research: Exploring its conceptualization and operationalization. Quality and Quantity, 52 (4), 1893–1907.
Moser, A., & Korstjens, I. (2018). Series: Practical guidance to qualitative research. Part 3: Sampling, data collection and analysis. European Journal of General Practice, 24 (1), 9–18.
Marlett, N., Shklarov, S., Marshall, D., Santana, M. J., & Wasylak, T. (2015). Building new roles and relationships in research: A model of patient engagement research. Quality of Life Research : an international journal of quality of life aspects of treatment, care and rehabilitation, 24 (5), 1057–1067.
Demian, M. N., Lam, N. N., Mac-Way, F., Sapir-Pichhadze, R., & Fernandez, N. (2017). Opportunities for engaging patients in kidney research. Canadian Journal of Kidney Health and Disease, 4 , 2054358117703070–2054358117703070.
Noyes, J., McLaughlin, L., Morgan, K., Roberts, A., Stephens, M., Bourne, J., Houlston, M., Houlston, J., Thomas, S., Rhys, R. G., et al. (2019). Designing a co-productive study to overcome known methodological challenges in organ donation research with bereaved family members. Health Expectations . 22(4):824–35.
Piil, K., Jarden, M., & Pii, K. H. (2019). Research agenda for life-threatening cancer. European Journal Cancer Care (Engl), 28 (1), e12935.
Hofmann, D., Ibrahim, F., Rose, D., Scott, D. L., Cope, A., Wykes, T., & Lempp, H. (2015). Expectations of new treatment in rheumatoid arthritis: Developing a patient-generated questionnaire. Health Expectations : an international journal of public participation in health care and health policy, 18 (5), 995–1008.
Jun, M., Manns, B., Laupacis, A., Manns, L., Rehal, B., Crowe, S., & Hemmelgarn, B. R. (2015). Assessing the extent to which current clinical research is consistent with patient priorities: A scoping review using a case study in patients on or nearing dialysis. Canadian Journal of Kidney Health and Disease, 2 , 35.
Elsie Baker, S., & Edwards, R. (2012). How many qualitative interviews is enough? In National Centre for Research Methods Review Paper . National Centre for Research Methods. http://eprints.ncrm.ac.uk/2273/4/how_many_interviews.pdf .
Sandelowski, M. (1995). Sample size in qualitative research. Research in Nursing & Health, 18 (2), 179–183.
Sim, J., Saunders, B., Waterfield, J., & Kingstone, T. (2018). Can sample size in qualitative research be determined a priori? International Journal of Social Research Methodology, 21 (5), 619–634.
Download references
Acknowledgements
no external funding.
Author information
Authors and affiliations.
Department of Neurology, Heidelberg University Hospital, Im Neuenheimer Feld 400, 69120, Heidelberg, Germany
Loraine Busetto, Wolfgang Wick & Christoph Gumbinger
Clinical Cooperation Unit Neuro-Oncology, German Cancer Research Center, Heidelberg, Germany
Wolfgang Wick
You can also search for this author in PubMed Google Scholar
Contributions
LB drafted the manuscript; WW and CG revised the manuscript; all authors approved the final versions.
Corresponding author
Correspondence to Loraine Busetto .
Ethics declarations
Ethics approval and consent to participate, consent for publication, competing interests.
The authors declare no competing interests.
Additional information
Publisher’s note.
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/ .
Reprints and permissions
About this article
Cite this article.
Busetto, L., Wick, W. & Gumbinger, C. How to use and assess qualitative research methods. Neurol. Res. Pract. 2 , 14 (2020). https://doi.org/10.1186/s42466-020-00059-z
Download citation
Received : 30 January 2020
Accepted : 22 April 2020
Published : 27 May 2020
DOI : https://doi.org/10.1186/s42466-020-00059-z
Share this article
Anyone you share the following link with will be able to read this content:
Sorry, a shareable link is not currently available for this article.
Provided by the Springer Nature SharedIt content-sharing initiative
- Qualitative research
- Mixed methods
- Quality assessment
Neurological Research and Practice
ISSN: 2524-3489
- Submission enquiries: Access here and click Contact Us
- General enquiries: [email protected]

Change Password
Your password must have 8 characters or more and contain 3 of the following:.
- a lower case character,
- an upper case character,
- a special character
Password Changed Successfully
Your password has been changed
- Sign in / Register
Request Username
Can't sign in? Forgot your username?
Enter your email address below and we will send you your username
If the address matches an existing account you will receive an email with instructions to retrieve your username
Mixed-Methods Design in Biology Education Research: Approach and Uses
- Abdi-Rizak M. Warfa
Search for more papers by this author
Educational research often requires mixing different research methodologies to strengthen findings, better contextualize or explain results, or minimize the weaknesses of a single method. This article provides practical guidelines on how to conduct such research in biology education, with a focus on mixed-methods research (MMR) that uses both quantitative and qualitative inquiries. Specifically, the paper provides an overview of mixed-methods design typologies most relevant in biology education research. It also discusses common methodological issues that may arise in mixed-methods studies and ways to address them. The paper concludes with recommendations on how to report and write about MMR.
INTRODUCTION
An increasing number of studies in biology education are reporting the use of mixed-methods research (MMR), in which quantitative and qualitative data are combined to investigate questions of interest in biology teaching and learning (e.g., Andrews et al. , 2012 ; Jensen et al. , 2012 ; Höst et al. , 2013 ; Ebert-May et al. , 2015 ; Seidel et al. , 2015 ). This increase coincides with general growth and expanded interest in mixed-methods approaches to research in various fields of study over the past 30 years ( Plano Clark, 2010 ). Consequently, several handbooks and articles have been written that describe the use of mixed methods in the social and behavioral sciences ( Tashakkori and Teddlie, 1998 , 2010 ; Creswell et al. , 2003 ; Creswell and Plano Clark, 2011 ; Greene, 2008 ; Terrell, 2011 ), focusing on both the theoretical underpinnings and procedural steps of conducting MMR. However, given the disciplinary ethos and divergent content perspectives of academic disciplines, it is important that researchers planning to use MMR become familiar with the theory and designs most commonly used within their disciplinary context. This article, therefore, focuses on the various ways in which quantitative and qualitative methods can be combined to address questions of interest in biology education and the many productive ways in which MMR can be used to support claims about biology teaching and learning.
The paper is organized into three parts. Part 1 provides introductory remarks that situate MMR within the larger context of research paradigms in science education. Part 2 provides a general description of mixed-methods approaches commonly found in biology education research (BER). Part 3 provides general guidelines on how to select an appropriate MMR design and attend to methodological issues that may arise when using MMR.
PART 1: UNDERSTANDING MIXED METHODS
Mixed methods emerged as a credible research design on the heels of a larger debate on research paradigms in education and the social sciences in the mid-1980s ( Johnson and Onwuegbuzie, 2004 ; Tashakkori and Teddlie, 2010 ; Treagust et al. , 2014 ). Biology researchers, however, have long used mixed-methods approaches to address issues of interest in biological sciences. It is, for example, common to determine the effect of a gene mutation by quantitative analysis and then characterize the context of that effect through qualitative analysis. It is also common to define behaviors of an animal and then count and analyze their frequency in different circumstances. In educational settings, the new approach provided a “third methodological” pathway that permitted combining quantitative and qualitative modes of social inquiry ( Johnson et al. , 2007 ; Tashakkori and Teddlie, 2010 ). In the words of Jennifer Greene (2008) ,
A mixed-methods way of thinking is an orientation toward social inquiry that actively invites us to participate in dialogue about multiple ways of seeing and hearing, multiple ways of making sense of the social world, and multiple standpoints on what is important and to be valued and cherished. (p. 20)
Green's description captures the essence of mixed methods—a pragmatic choice to address research problems through multiple methods with the goal of increasing the breadth, depth, and consistency of research findings. Integration of research findings from quantitative and qualitative inquiries in the same study or across studies maximizes the affordances of each approach and can provide better understanding of biology teaching and learning than either approach alone. While quantitative methods can reveal empirical evidence showing causal or correlative relationships or the effects of interventional studies, qualitative methods provide contextual information that colors the experiences of individual learners. The goal of mixed methods is not, however, to replace either the quantitative or the qualitative approaches. Certain problems—for example, addressing gains in standardized test scores—are better addressed through quantitative methods (e.g., Knight and Wood, 2005 ), and some—for example, understanding the meaning students assign to reaction arrows—merit qualitative research (e.g., Wright et al ., 2014 ). Rather, the goal of mixed methods is to build on the strengths of both methods and minimize their weaknesses when the research merits using more than one method ( Creswell et al. , 2003 ; Johnson and Onwuegbuzie, 2004 ). Recent studies from the biology education literature will help illustrate the types of research that benefit from a mixed-methods approach.
In a recent study that used both quantitative and qualitative methods, Seidel et al. (2015) investigated non–content related conversational language, such as procedural talk, used by course instructors in a large reform-based introductory biology classroom cotaught by two instructors. Such language, which the authors termed “Instructor Talk,” is the language used to facilitate overall learning in the classroom, for example, language used to give directions on homework assignments or justifying use of active-learning strategies. Instructor Talk is distinct from language used to describe specific course concepts. To understand the prevalence of such language in biology classrooms, the authors asked, “What types of Instructor Talk exist in a selected introductory college biology course?” This question was exploratory in nature and merited qualitative inquiry that focused on identifying the types of Instructor Talk the two instructors used. The authors’ subsequent question, “To what extent do two instructors differ in the types and quantity of Instructor Talk they appear to use?,” aimed to enhance the findings from the qualitative phase and provided ways to further study and generalize this construct in a variety of class types ( Seidel et al. , 2015 ). The authors were able to address their initial research question through analysis of classroom transcripts containing more than 600 instructor quotes, identifying five emergent categories that were present in the analyzed sessions. They followed this exploratory qualitative phase of the study with statistical analyses that compared how often the instructors used identified categories and the average instances of Instructor Talk per class session. Without first characterizing and identifying patterns of Instructor Talk through the exploratory initial qualitative data, the authors could not have addressed the second question. Neither qualitative nor quantitative method was sufficient to address both research questions, but combining them strengthened the overall findings of the study.
In another BER study, Andrews et al. (2012) used a mixed-methods study to investigate undergraduate biology students’ misconceptions about genetic drift. Using qualitative data analysis, the authors identified 16 misconceptions students held about genetic drift that fit into one of five broad categories (e.g., novice genetics, genetic drift comprehension). Subsequent use of quantitative methods examined the frequency of misconceptions present before and after introductory instruction on genetic drift. The quantitative data supplemented the results of the qualitative analysis and shed light on changes in student misconceptions as a result of instruction. In this study, although data collection was separated in time and space, the quantitative and qualitative analyses were integrated, and the different data sets were used to generate the categories of misconceptions about genetic drift and to corroborate the findings. Again, we see the utility of both methods within the same study.
The Andrews et al . (2012) and Seidel et al . (2015) studies illustrate the types of research problems that merit a mixed-methods study: research problems in which a single method, qualitative or quantitative, is insufficient to fully understand the problem ( Creswell et al ., 2003 ). Another role of MMR research is to use qualitative work to follow up/elaborate on quantitative findings or to validate findings in multiple ways. More broadly, teaching and learning occur in social environments with specific cultural contexts, personal value systems, and classroom dynamics that color how students learn and teachers teach. In such environments, understanding the educational processes in which teachers and students engage becomes crucial to understanding how students learn. MMR is particularly appropriate for BER, because it contextualizes quantitative differences observed in BER studies, capturing the contextual, sociocultural norms and the experiential factors that characterize undergraduate biology classrooms.
PART 2: GENERAL TYPOLOGIES OF MIXED-METHODS DESIGNS
For those questions that merit a mixed-methods approach, this section of the paper describes different typologies of mixed-methods designs available for biology educators. General guidelines on the use of MMR and the methodological issues to consider are described in Part 3.
Based on a review of the literature (see Creswell et al. , 2003 ), there are three general approaches to mixed methods—sequential, concurrent, or data transformation—that are most applicable to BER studies. These basic designs can get more complicated and advanced as merited by the phenomenon studied. More advanced variants of MMR, for example, multiphase designs or the transformative designs appropriate for social justice, are not addressed in this paper ( Creswell et al. , 2003 ; Terrell, 2011 ). The following paragraphs discuss each of the basic designs with respect to data-collection sequencing, method priority, and data-integration steps. As discussed below, these decisions often influence which MMR design to choose.
Sequential Designs
The sequential design approach implies linear data collection and analysis in which the collection of one set of data (e.g., qualitative) is followed by the collection and analysis of the other (e.g., quantitative). There are two general approaches within this design (see Figure 1, A and B ) based on the implementation sequence of the data and their intended usage ( Creswell et al. , 2003 ): 1) sequential explanatory and 2) sequential exploratory. Each subdesign, along with illustrative examples in biology education, is further described in the following sections.

FIGURE 1. Basic typologies of MMR. There are three basic designs of mixed methods that differ in how data collection and analysis is sequenced: (A) sequential explanatory design, in which the quantitative method precedes the qualitative method; (B) sequential exploratory design, in which the qualitative method precedes the quantitative method; and (C) concurrent triangulation design, in which qualitative and quantitative data are collected concurrently.
Sequential Explanatory Design
The sequential explanatory approach is characterized by two distinct phases: an initial phase of quantitative data collection and analysis followed by a second qualitative data-collection and analysis phase (see Figure 1A ). Findings from both phases are integrated during the data-interpretation stage. The general aim of this approach is to further explain the phenomenon under study qualitatively or to explore the findings of the quantitative study in more depth ( Tashakkori and Teddlie, 2010 ). Given the sequential nature of data collection and analysis, a fundamental research question in a study using this design asks, “In what ways do the qualitative findings explain the quantitative results?” ( Creswell et al. , 2003 ). Often, the initial quantitative phase has greater priority over the second, qualitative phase. At the interpretation stage, the results of the qualitative data often provide a better understanding of the research problem than simply using the quantitative study alone. As such, the findings in the quantitative study frequently guide the formation of research questions addressed in the qualitative phase ( Creswell et al. , 2003 ), for example, by helping formulate appropriate follow-up questions to ask during individual or focus group interviews. The following examples from the extant literature illustrate how this design has been used in the BER field.
In an interventional study with an overtly described two-phase sequential explanatory design, Buchwitz et al. (2012) assessed the effectiveness of the University of Washington’s Biology Fellows Program, a premajors’ course that introduced incoming biology majors to the rigor expected of bioscience majors and assisted them in their development as science learners. The program emphasized the development of process skills (i.e., data analysis, experimental design, and scientific communication) and provided supplementary instruction for those enrolled in introductory biology courses. To assess the effectiveness of the program, the authors initially used nonhierarchical linear regression analysis with six explanatory variables inclusive of college entry data (high school grade point average and Scholastic Aptitude Test scores), university-related factors (e.g., economically disadvantaged and first-generation college student status), program-related data (e.g., project participation), and subsequent performance in introductory biology courses. Analysis showed that participation in the Biology Fellows Program was associated with higher grades in two subsequent gateway biology courses across multiple quarters and instructors. To better understand how participating in the Biology Fellows Program may be facilitating change, the authors asked two external reviewers to conduct a focus group study with program participants. Their goal was to gather information from participants retrospectively (2 to 4 years after their participation in the program) about their learning experiences in and beyond the program and how those experiences reflected program goals. Students’ responses in the focus group study were used to generate themes and help explain the quantitative results. The manner in which the quantitative and qualitative data were collected and analyzed was described in detail. The authors justified the use of this design by stating, “A mixed-methods approach with complementary quantitative and qualitative assessments provides a means to address [their research] question and to capture more fully the richness of individuals’ learning” (p. 274).
In a similar study, Fulop and Tanner (2012) administered written assessments to 339 high school students in an urban school district and subsequently interviewed 15 of the students. The goal of this two-phased sequential study was to examine high school students’ conceptions about the biological basis of learning. To address their research problem, they used two questions to guide their study: 1) “After [their] mandatory biology education, how do high school students conceptualize learning?,” and 2) “To what extent do high school students have a biological framework for conceptualizing learning?” The authors used statistical analysis (post hoc quantitative analysis and quantification of open-ended items) to score the written assessment and used thematic analysis to interpret the qualitative data. Although the particular design of the sequential explanatory model is not mentioned in the article, the authors make it clear that they used a mixed-methods approach and particularly mention how the individual interviews with a subset of students drawn from the larger study population were used to further explore how individual students think about learning and the brain. In drawing their conclusions about students’ conceptualization of the biological basis of learning, the authors integrated analysis of the quantitative and qualitative data. For example, on the basis of the written assessment, the authors concluded that 75% of the study participants demonstrated a nonbiological framework for learning but also determined that 67% displayed misconceptions about the biological basis of learning during the interviews.
Sequential Exploratory Design
The sequential exploratory approach is similarly characterized by two distinct phases: an initial qualitative phase followed by a second phase of quantitative data collection and analysis (see Figure 1B ). Similar to the sequential explanatory approach, findings from both phases in this design are integrated during the data-interpretation stage. Unlike the sequential explanatory approach, the general aim of this approach is to further explore the phenomenon under study quantitatively or to perform quantitative studies to generalize qualitative findings to different samples ( Tashakkori and Teddlie, 2010 ). Given the sequential nature of data collection and analysis, a fundamental research question in a study using this design often asks, “In what ways do the quantitative findings generalize the qualitative results?” ( Creswell et al. , 2003 ).
As a research method, the sequential exploratory approach is often the most appropriate design when developing new instruments or when a researcher intends to generalize findings from a qualitative study to different groups ( Tashakkori and Teddlie, 1998 , 2010 ; Creswell et al. , 2003 ). Consider, for example, the case of a biology education researcher interested in examining student misconceptions in evolution. Using the sequential exploratory approach, the researcher would collect qualitative data from interviews to identify commonly held student misconceptions in evolutionary concepts. The researcher can then use the qualitative data to develop an instrument on evolution misconceptions that allows the collection of quantitative data from a large number of participants in various settings and institutions (after instrument validation and psychometric analysis). In this case, the initial qualitative data would inform the design of the instrument used to collect the quantitative data, often using identified student misconceptions as distractors. An example of studies that followed the instrument development process outlined here can be found in Hanauer and Dolan (2013) and Hanauer et al. (2012) .
Pugh et al . (2014) used the sequential exploratory design in a study that investigated high school biology students’ conceptual understanding of the concept of natural selection and their ability to generatively use the newly learned concepts across knowledge domains in biology. To assess students’ transfer ability and conceptual understanding, the authors first collected qualitative data by administering open-response items to 138 students and were able to identify, on the basis of thematic analysis, particular patterns of surface and deep-level transfer. Subsequently, the authors collected quantitative data that showed a small but significant relationship between deep-level, but not surface-level, transfer and conceptual understanding. The principal methodology of the study was qualitative in nature and in turn informed the quantitative component of the study. The combination of the two methods shed light on the relationship between concept understanding and the patterns of knowledge transfer.
Strengths and Weaknesses of the Sequential Designs
In both of the sequential models described above (exploratory and explanatory), the data collection and analysis proceeded in two distinct phases. As illustrated by the examples from the BER literature, the main strengths of the sequential designs include the ability to 1) contextualize and generalize qualitative findings to larger samples (in the case of sequential exploratory); 2) enable one to gain a deeper understanding of findings revealed by quantitative studies (in the case of sequential explanatory); and 3) collect and analyze the different methods separately. Additionally, the two-phase approach makes sequential designs easy to implement, describe, and report.
One weakness of sequential designs is the length of time required to complete both data-collection phases, especially given that the second phase is often in response to the results of the first phase. That is, by collecting the data at two different time points, one essentially doubles the length of time required to complete a single-method study. Moreover, because data collection is sequential, it may be difficult to decide when to proceed to the next phase. It may also be difficult to integrate or connect the findings of the two phases. For those projects with shorter time lengths, concurrent designs in which both data sets are collected in a single phase may be more appropriate. The next section of the paper provides details of concurrent designs of MMR.
Concurrent Designs
In the concurrent design, both qualitative and quantitative data are collected in a single phase. Because the general aim of this approach is to better understand or obtain more developed understanding of the phenomenon under study, the data can be collected from the same participants or similar target populations. The goal is to obtain different but complementary data that validate the overall results. There are two basic approaches within concurrent design: 1) concurrent triangulation ( Figure 1C ) and 2) concurrent nested ( Figure 2A ). These are described below.

FIGURE 2. Complex typologies of MMR. Two complex forms of MMR are (A) concurrent nested design, in which either the qualitative or the quantitative method is nested within a primary quantitative or qualitative approach (in this case, the main difference hinges on data prioritization); and (B) transformation design, in which one data form is transformed to the other (e.g., qualitative to quantitative).
Concurrent Triangulation Design
The concurrent triangulation design is the most common approach used in BER studies. The main objective is to corroborate or cross-validate findings by using both quantitative and qualitative studies. Data collection and analysis is done separately but merged afterward (see Figure 1C ). In interpreting the overall merged results ( Figure 1C ), one looks for data convergence, divergence, contradictions, or any relationship the separate data analyses reveal. This can be done using several strategies, for example, through side-by-side comparison that discusses how the findings of one data set confirm or refute findings of the other data set. As the following examples from the BER literature illustrate, one method (qualitative or quantitative) can have priority over the other in the concurrent triangulation approach.
In a recent study that used the concurrent triangulation approach, Jensen et al. (2012) explored the effectiveness of a first-year class project in supporting student progress toward selected student learning and development outcomes. The students were required to complete a group video project focused on nutrition and healthy eating as a capstone class assignment. Using a structured rubric to track frequency data, the authors collected and analyzed quantitative measures of student behavior. They similarly collected qualitative data through observations and interviews with representative individual students and a focus group. In justifying why they used this design, the authors stated, “the data-collection techniques used in this study provide a degree of triangulation aimed at establishing validity of the conclusions drawn from the evaluation” (p. 72). The study was primarily qualitative in nature, with the objective of understanding a particular student experience validated by the quantitative measures. In this case, the authors found convergent results that strengthened the overall study—student behaviors as measured by both the quantitative and qualitative results were consistent with targeted learning and development outcomes.
In a similar manner, Höst et al. (2013) used a concurrent triangulation design to investigate the impact of using two external representations of virus self-assembly, an interactive tangible three-dimensional model or a static two-dimensional image, on student learning about the process of viral self-assembly. All the students in a biochemistry course at a Swedish university engaged in a small-group exercise that included the same series of tasks. They were randomly assigned to groups, some of which used the three-dimensional tangible model and some of which used the static images. Students completed a test before and after the group exercise. Researchers used an analysis of variance to test for an association between two factors (external representation: tangible model versus image; and testing time: pre versus post) and the score a student earned on the test. The authors found that test scores differed between the pre- and posttests but not between the two types of external representations. The researchers used qualitative analyses of open-response questions to further assess how the group exercise influenced students’ conceptual understanding of self-assembly. The findings from the qualitative analysis corroborated the findings from the quantitative analysis.
In the preceding examples of the concurrent triangulation design, the authors collected quantitative and qualitative data concurrently, using closed-ended and open-ended items. In the Jensen et al. (2012) study, the qualitative data had priority over the quantitative data. In the Höst et al. (2013) study, the quantitative data appear to have had priority over the qualitative data. In concurrent triangulation studies, either method can have priority over the other or both can be on equal footing. In both studies, the authors justified their use of the specific concurrent method as a way to triangulate their findings. The next section contrasts this triangulation approach with the concurrent nested design.
Concurrent Nested Design
In the concurrent nested design, a strong supplemental study is collected during the data-collection and analysis phase of the primary study (see Figure 2A ). In this type of study, it does not matter whether the primary study is quantitative and the nested study qualitative or vice versa. The major aim of this design, which is often used in the health sciences, is to use the nested analysis to address different research questions than those addressed by the primary method or to use the nested method to seek information about different levels of the research problem. The general idea is that a need arises to address different types of questions within the research project that require different methods ( Creswell et al. , 2003 ). Most published mixed-methods studies that use this approach tend to be experimental designs that require qualitative aspects to examine how an intervention is working or to follow up specific aspects of the experiments ( Creswell et al. , 2003 ). An illustrative example in the context of biology education would be for a researcher to implement a specific interventional study in his or her classroom and to evaluate the effectiveness of the intervention by using quasi-experimental pre/posttest measures. However, in between the two measurements, the researcher interviews or collects open-ended written responses from the students to examine students’ experiences with the intervention.
A study by Tomanek and Montplaisir (2004) used this approach. In this study, the authors examined the study habits of students enrolled in a large introductory biology class. The authors collected two types of data: 1) pre/posttest assessment data from lecture sessions that covered cell division and Mendelian genetics, and 2) preinstruction and postinstruction interviews with a purposely selected sample of 13 students. The pre/posttest items constituted the quantitative data, while the pre/postinstruction interviews constituted the qualitative data. Because the main goal of the study was to understand students’ study habits both during lecture (e.g., how they used the information presented in lectures) and outside the classroom (e.g., how they studied, what resources they used), the study was primarily qualitative. The quantitative data were concurrently collected but addressed a different question: Did the study tasks and habits in which the students engaged help them academically? The quantitative data were thus nested within the larger qualitative study, illustrating the general scheme of the concurrent nested design. This research illustrates a distinguishing feature of nested and triangulated designs: in the nested design the two methods of analysis often address different research questions, whereas in triangulated design the two methods address the same question.
Strengths and Weaknesses of the Concurrent Designs
In both of the concurrent designs described above (triangulation and nested), the data collection occurs during a single phase of the research and the analysis occurs separately. Given the shorter period of time and the separate nature of the analysis, the concurrent designs tend to be the most efficient of the mixed-methods typology. There are two main drawbacks of the concurrent designs. For one, the concurrent nature of data collection precludes follow-up on any interesting or confusing issues that may arise as analysis unfolds. Second, data integration may become an impediment if the results are contradictory and/or diverge. The difficulty in this case becomes how to resolve divergent results short of declaring the study a failure. Additionally, it may be difficult to compare and contrast qualitative and quantitative data without transforming them to a common scale—for example, by transforming the qualitative data to dichotomous variables that can be subjected to statistical analysis, thus enabling comparison with the other quantitative data. However, as described later, such transformation may sacrifice the depth and the contextual data associated with the qualitative research.
Data-Transformation Designs
The data-transformation design implies changing one data set (e.g., qualitative) to another (e.g., quantitative) through either quantitating or qualitizing . Quantitating refers to the act of transforming qualitative data (codes) into quantitative data (variables), whereas qualitizing is the act of transforming numerical data (variables) into codes (or themes) that can be analyzed qualitatively ( Tashakkori and Teddlie, 2010 ). For example, Witcher et al. (2001) examined preservice teachers’ perceptions of the characteristics of effective teachers. To address their research questions, the authors collected quantitative data but then transformed those data into six general themes (e.g., student-centeredness, enthusiasm) that were prevalent in the participants’ responses. While qualitizing quantitative data, as done by Witcher et al. (2001) , is theoretically possible, in practice, quantitating of qualitative data is far more common, a practice reinforced by the rhetorical appeal of numbers and their association with rigor ( Sandelowski et al. , 2009 ). Figure 2B shows a general scheme of qualitative data that are transformed into quantitative form. As can be seen in the figure, data collection can happen sequentially or concurrently (as described earlier) for the sequential and concurrent designs. The underlying rationale for choosing this design is also similar to that described previously for sequential and concurrent designs. The only difference in this case is that one data set (quantitative or qualitative) is transformed. Comparison and merging of the two data sets occurs at the data-interpretation stage.
The quantitative survey data were entered into a chosen database (e.g., Excel) and organized under two broad categories of positive and negative attitudes.
The qualitative interview data were also analyzed and coded as positive and negative responses. This data set was then quantified into dichotomous variables 0 or 1 based on the absence or presence of negative and positive responses.
The two data sets were merged, and the combined data were analyzed for association using statistical analyses.
The overall data interpretation examined the prevalence of positive versus negative attitudes in the student population.
In this hypothetical case, the qualitative interview data are transformed into dichotomous variables corresponding to negative and positive attitudinal aspects of CUREs, categories predetermined before data collection. The overall data analysis occurred after the qualitative and the quantitative data were integrated. The goal of the interview data was to capture any contextual variables that were not explained by the survey data (not to triangulate the findings). That is, the interview data had been used to interpret the survey data and fill any holes that the survey did not capture, as it might not have identified a priori all the things that students might have feelings about with respect to CURE labs. The combined data thus provided a more complete picture than was possible only with the survey (i.e., quantitative) data.
The study by Ebert-May et al. (2015) is also an example of this type of research. In their study, Ebert-May and colleagues examined the extent to which biology postdoctoral fellows (postdocs) believed in and implemented evidence-based pedagogies after completion of a 2-year professional development program. The authors used the Approaches to Teaching Inventory ( Trigwell and Prosser, 2004 ) and local surveys to characterize the postdocs’ beliefs about teaching and knowledge and experiences with active-learning pedagogies. To capture and analyze the postdocs’ teaching practices, the participants submitted videos for at least two complete class sessions for each full course that they taught while participating in the professional development program. To analyze teaching practices captured by the videos, the authors chose a validated measurement of teacher practices in the classroom, Reformed Teaching Observation Protocol ( Sawada et al. , 2002 ), to measure the degree to which classroom instruction used active-learning pedagogies. The authors did not develop thematic analysis of the postdocs’ teaching practices but rather transformed the qualitative video data into numerical units that were analyzed with statistical tools. Thus, despite the collection of both quantitative data (i.e., surveys) and qualitative data (i.e., videos), only quantitative data-analysis strategies were ultimately used to examine their research question.
Strengths and Weaknesses of the Transformation Designs
The transformation designs can enable researchers to convert qualitative data into a quantitative format to meet specific goals of quantitatively oriented research, such as evaluating the effect size of an intervention or generalizing. This advantage, however, also reveals a major weakness of this design: transformation of qualitative into quantitative data causes some richness and depth of the qualitative data to be lost. Some researchers (e.g., Bazeley, 2004 ) contend that transforming qualitative data into dichotomous variables makes them one-dimensional and strips them of the flexibility associated with thematic analysis; that is, the quantitated data are no longer mutable to analysis of emergent themes characteristic of qualitative research. For this reason, the transformation designs may be most effective when the focus is quantification of a phenomenon rather than an in-depth, comprehensive understanding of the phenomenon.
PART 3: PRACTICAL GUIDELINES AND ISSUES TO CONSIDER
Having determined that a research question merits a mixed-methods approach, it is necessary to select an appropriate MMR design. As we have seen, the major influence on which design to choose is driven by data-collection sequencing, method priority, and the planned data-integration steps. Figure 3 provides summary guidelines on how to select a specific design among the different MMR topologies discussed in the previous section. In this section, we discuss major methodological issues that may arise during the study design.

FIGURE 3. MMR design decision tree. This if–use dichotomous key is designed to help researchers select appropriate MMR design based on the intent of their research. Refer to part 2 for the main differences between the different MMR designs.
Methodological Issues to Consider
In addition to issues surrounding the selection of an appropriate MMR design, several methodological issues such as those listed in Table 1 may arise. These issues (sampling, participant burden, data analysis, and transparency) are crucial in all research methods but more so when mixing quantitative and qualitative methods. For example, if a researcher collects both qualitative and quantitative data from the same participants, what burden does that place on the participants? On the other hand, if the researcher collects data from different participants, what complications does that present for data analysis? One has to be cognizant of both the burden and the complexity that arises from data collection when conducting a mixed-methods study.
| Issues to consider | Questions to ask |
|---|---|
| 1. Sample size | |
| ○ Sample size is likely to vary between quantitative and qualitative work due to practical concerns like money, time, effort, and the main purposes of the study. | |
| ○ A targeted or deliberate sample often better meets study objectives in qualitative work, whereas a large sample is necessary for quantitative work. One must consider trade-offs in sample size when deciding which method should take priority in the chosen design | |
| 2. Participant burden | |
| ○ If collecting both qualitative and quantitative data, do both kinds of data need to be collected from the same individuals? | |
| ○ A researcher must consider what issues could arise if data are collected from different participants | |
| 3. Analysis and interpretative issues | |
| ○ One must consider how analysis of different types of data can strengthen the collection and analysis of the other data type, and plan the study accordingly | |
| ○ One must also recognize that study design may need to change midstudy based on early findings. | |
| 4. Transparency | |
| ○ For each design, one must report how the mixed-methods study design addressed shortcomings often associated with single-method studies | |
| ○ For sequential designs, how did the results in the first phase inform research processes in the second phase? | |
| ○ For concurrent designs, how was data integrated, especially if the findings diverged? | |
| ○ For data-transformation designs, was richness and depth that was associated with the qualitative approach lost when data were transformed? If so, does this loss affect study outcomes? |
Sampling is one issue that may present complications, as it is likely to vary between quantitative and qualitative work due to practical concerns like money, time, and effort, but also because the purpose of these different methods varies. The purpose of qualitative work is generally not to infer to a broader population, so a large random sample is not necessary, as may be the case in a quantitative study. The trick for a researcher designing a mixed-methods study is to consider questions that are specific to quantitative analyses (i.e., power) and those that are specific to qualitative analysis (i.e., variations in views and perspectives, representativeness of the qualitative sample). Thus, one has to consider what sample size is appropriate to make useful inferences from both the quantitative and qualitative data.
Certain sampling issues are specific to mixed-methods approaches, For example, one must consider whether the interview sample must be a subset of the quantitative sample or from a different population. If it is a subset, how will that subset be targeted—for example, should it be representative of extreme performances or large enough to capture the most common groups? There are advantages and disadvantages to both approaches. If one chooses to represent extreme performances, that allows using a smaller sample but necessitates a sequential or at least a nested design. If, on the other hand, one chooses to interview a sample large enough to capture the most common groups, trade-offs come in terms of investment of time and resources versus the ability to make generalizable inferences as a result of the large sample size. These issues require thought and attention as one designs a mixed-methods study.
Data integration is another methodological issue that may arise. For example, in sequential designs, data analysis occurs separately, and findings are integrated at the interpretation stage. In contrast, when using concurrent triangulation designs, data analysis occurs simultaneously. Theoretically, data analysis can occur at any point in the research process. So, when is the best time for analyzing the data and how should they be integrated? Some authors (e.g., Yoshikawa et al. , 2008 ) argue that it is not the best methodological choice to separate analysis of quantitative and qualitative data but instead it is preferable to integrate the results throughout the analysis phase of the research project. Yoshikawa et al. (2008) argue that such an integration approach (i.e., integrating the results throughout the analysis phase) can result in rich integration across methods and analyses. Such integration, however, will require expertise and a certain level of competence in both qualitative and quantitative methods, as qualitative and quantitative methods require different skill sets. These issues can be resolved through collaboration and utilization of the skill sets of different team members who can take the lead in specific aspects of the research while communicating with the research team on the results.
Finally, given the varied nature and purposes of the different methods used in MMR, it is important to report in detail how analyses are conducted. For sequential designs, the need is to discuss matters such as how results from the first phase informed data collection and analysis in the second phase. For concurrent designs, the need is to discuss what strategies are used to resolve conflicts that may arise from contradictory results (e.g., gathering more data to address the conflict). One must also describe how it was ensured that the depth and flexibility associated with qualitative data were not lost in the analysis, especially if a transformation design was used. If at all possible, it helps to publish data-collection tools such as interview protocols, quantitative instruments, and visual representations of data-integration plans. That is, methodological transparency becomes an important consideration in mixed-methods studies.
Recommendations for Writing about MMR
communicate the intent of the study (e.g., “this mixed-methods study examined …”);
specify which design was used (e.g., “we used sequential exploratory design to…”);
describe how both data forms were collected (e.g., “through structured interviews , we addressed the question of … participants were also surveyed …”);
provide the rationale for why both quantitative and qualitative data sets were collected (e.g., “the qualitative study addressed [i.e., the research question]; the quantitative study addressed [i.e., the research question]…”); and
describe how validity and reliability (or “trustworthiness”) were established in the chosen design.
Ivankova et al . (2006) provide a sample MMR study featuring most of the elements outlined above and discuss additional guidelines on how to communicate about MMR studies. Table 2 provides further references on general approaches to MMR and writing about mixed methods, including the use of diagrams. Mixed methodologists particularly recommend the use of visual representations to depict the procedural steps involved in a mixed-methods study, such as the one shown in Figure 4 for a hypothetical two-phased sequential explanatory study. As can be seen in Figure 4 , the visual diagram depicts the progression of research activities from data collection and analysis in the initial quantitative phase to qualitative data collection and analysis in the second phase to questions that may help integrate and interpret the findings. Most studies in the MMR literature use similar visual depictions that portray the complexity and the sequence of research activities in MMR studies ( Ivankova et al. , 2006 ; Creswell and Plano Clark, 2011 ).

FIGURE 4. Sample diagram for a hypothetical BER study. Researchers using mixed-methods approaches are encouraged to visually depict the procedural steps involved in their study, as shown in this figure.
| Key references on MMR (general approaches and methodological issues) | . Designing and Conducting Mixed Methods Research, 2nd ed., Thousand Oaks, CA: Sage |
| . Paradigms lost and pragmatism regained: methodological implications of combining qualitative and quantitative methods. Journal of Mixed Methods Research 1, 48–76. | |
| . Mixed Methodology: Combining Qualitative and Quantitative Approaches, Thousand Oaks, CA: Sage. | |
| . SAGE Handbook of Mixed Methods in Social and Behavioral Research, 2nd ed., Thousand Oaks, CA: Sage. | |
| References for providing justification for mixed-methods studies | . Integrating quantitative and qualitative research: how is it done? Qualitative Research 6, 97–113. |
| . Is mixed methods social inquiry a distinctive methodology? Journal of Mixed Methods Research 2, 7–22. | |
| . Mixed methods research: a research paradigm whose time has come. Educational Researcher 33(7), 14–26. | |
| References for how to write about MMR | . Designing and Conducting Mixed Methods Research, 2nd ed., Thousand Oaks, CA: Sage. |
| ., (2006). Using mixed methods sequential explanatory design: from theory to practice. Field Methods 18, 3–20. | |
| . Research questions in mixed methods research. In: Handbook of Mixed Methods Research, 2nd ed., ed. A. Tashakkori and C. Teddlie, Thousand Oaks, CA: Sage, 275–304. |
In the realm of biology education, the social nature of educational inquiry often merits the use of multiple perspectives, as was the case with the BER studies cited in this paper. Given the various approaches of mixed methods discussed here and elsewhere ( Creswell et al. , 2003 ; Tashakkori and Teddlie, 2010 ), it is important for mixed-methods researchers to describe the decisions that went into their MMR design selections and guided their research projects. Three factors were discussed in this paper that can guide that selection: data-collection sequencing, method priority, and data integration. Understanding how these factors effect which MMR design to select, being clear about data-analysis procedures, and attending to methodological issues that arise will only strengthen MMR studies in our field and undoubtedly enrich biology teaching and learning through the use of multiple perspectives.
ACKNOWLEDGMENTS
I thank Drs. Anita Schuchardt and James Nyachwaya for close reading of this article and the editor and two anonymous reviewers for their constructive feedback on an earlier version of this article.
- Andrews TM, Price RM, Mead LS, McElhinny TL, Thanukos A, Perez KE, Herreid CF, Terry DR, Lemons PP ( 2012 ). Biology undergraduates’ misconceptions about genetic drift . CBE Life Sci Educ 11 , 248-259. Link , Google Scholar
- Bazeley P ( 2004 , Ed. R BuberJ GadnerL Richards , Issues in mixing qualitative and quantitative approaches to research In: Applying Qualitative Methods to Marketing Management Research , London, UK: Palgrave Macmillan, 141-156. Google Scholar
- Bryman A ( 2006 ). Integrating quantitative and qualitative research: how is it done? . Qual Res 6 , 97-113. Google Scholar
- Buchwitz BJ, Beyer CH, Peterson JE, Pitre E, Lalic N, Sampson PD, Wakimoto BT ( 2012 ). Facilitating long-term changes in student approaches to learning science . CBE Life Sci Educ 11 , 273-282. Link , Google Scholar
- Creswell JW, Plano Clark VL ( 2011 ). Designing and Conducting Mixed Methods Research , 2nd ed. Thousand Oaks, CA: Sage. Google Scholar
- Creswell JW, Plano Clark VL, Guttmann ML, Hanson WE ( 2003 , Ed. A TashakkoriC Teddlie , Advanced mixed methods research designs In: Handbook of Mixed Methods in Social and Behavioral Research , Thousand Oaks, CA: Sage, 209-240. Google Scholar
- Ebert-May D, Dertig TL, Henkel TP, Maher JM, Momsen JL, Arnold B, Passmore HA ( 2015 ). Breaking the cycle: future faculty begin teaching with learner-centered strategies after professional development . CBE Life Sci Educ 14 , ar22. Link , Google Scholar
- Fulop RM, Tanner KD ( 2012 ). Investigating high school students’ conceptualization of the biological basis of learning . Adv Physiol Educ 36 , 131-142. Medline , Google Scholar
- Greene JC ( 2008 ). Is mixed methods social inquiry a distinctive methodology? . J Mixed Methods Res 2 , 7-22. Google Scholar
- Hanauer DI, Dolan EL ( 2013 ). The project ownership survey: measuring differences in scientific inquiry experiences . CBE Life Sci Educ 13 , 149-158. Google Scholar
- Hanauer DI, Frederick J, Fotinakes B, Strobel SA ( 2012 ). Linguistic analysis of project ownership for undergraduate research experiences . CBE Life Sci Educ 11 , 378-385. Link , Google Scholar
- Höst GE, Larsson C, Olson A, Tibell LAE ( 2013 ). Student learning about biomolecular self-assembly using two different external representations . CBE Life Sci Educ 12 , 471-482. Link , Google Scholar
- Ivankova NV, Creswell JW, Stick S ( 2006 ). Using mixed methods sequential explanatory design: from theory to practice . Field Methods 18 , 3-20. Google Scholar
- Jensen M, Mattheis A, Johnson B ( 2012 ). Using student learning and development outcomes to evaluate a first-year undergraduate group video project . CBE Life Sci Educ 12 , 68-80. Google Scholar
- Johnson RB, Onwuegbuzie AT ( 2004 ). Mixed methods research: a research paradigm whose time has come . Educ Res 33 , (7), 14-26. Google Scholar
- Johnson RB, Onwuegbuzie AT, Tuner LA ( 2007 ). Toward a definition of mixed methods research . J Mixed Methods Res 1 , 112-133. Google Scholar
- Knight JK, Wood WB ( 2005 ). Teaching more by lecturing less . Cell Biol Educ 4 , 298-310. Link , Google Scholar
- Morgan DL ( 2007 ). Paradigms lost and pragmatism regained: methodological implications of combining qualitative and quantitative methods . J Mixed Methods Res 1 , 48-76. Google Scholar
- Plano Clark VL ( 2010 ). The adoption and practice of mixed methods: U.S. trends in federally funded health-related research . Qual Inq 16 , 428-440. Google Scholar
- Plano Clark VL, Badiee M ( 2010 , Ed. A TashakkoriC Teddlie , Research questions in mixed methods research In: Handbook of Mixed Methods Research , 2nd ed. Thousand Oaks, CA: Sage, 275-304. Google Scholar
- Pugh K, Koskey K, Linnenbrink-Garcia L ( 2014 ). High school biology students’ transfer of the concept of natural selection: a mixed-methods approach . J Biol Educ 48 , 23-33. Google Scholar
- Sandelowski M, Voils CI, Knalf G ( 2009 ). On quantitizing . J Mixed Methods Res 3 , 208-222. Medline , Google Scholar
- Sawada D, Piburn MD, Judson E, Turley J, Falconer K, Benford R, Bloom I ( 2002 ). Measuring reform practices in science and mathematics classrooms: the Reformed Teaching Observation Protocol . School Sci Math 102 , 245-253. Google Scholar
- Seidel SB, Reggi AL, Schinske JN, Burrus LW, Tanner KD ( 2015 ). Beyond the biology: a systematic investigation of noncontent Instructor Talk in an introductory biology course . CBE Life Sci Educ 14 , ar43. Link , Google Scholar
- Tashakkori A, Teddlie C ( 1998 ). Mixed Methodology: Combining Qualitative and Quantitative Approaches , Thousand Oaks, CA: Sage. Google Scholar
- Tashakkori A, Teddlie C ( 2010 ). SAGE Handbook of Mixed Methods in Social and Behavioral Research , 2nd ed. Thousand Oaks, CA: Sage. Google Scholar
- Terrell S ( 2011 ). Mixed-methods research methodologies . Qual Rep 17 , 254-280. Google Scholar
- Tomanek D, Montplaisir L ( 2004 ). Students’ studying and approaches to learning in introductory biology . Cell Biol Educ 3 , 253-262. Link , Google Scholar
- Treagust DF, Won M, Duit R ( 2014 , Ed. Norman LedermanSandra Abell , Paradigms in science education research In: Handbook of Research on Science Education, vol. 2 , 2nd ed. New York: Routledge, 3-17. Google Scholar
- Trigwell K, Prosser M ( 2004 ). Development and use of the approaches to teaching inventory . Educ Psychol Rev 16 , 409-424. Google Scholar
- Witcher A, Onwuegbuzie AJ, Minor LC ( 2001 ). Characteristics of effective teachers: perceptions of preservice teachers . Res Schools 8 , 45-57. Google Scholar
- Wright LK, Fisk JN, Newman DL ( 2014 ). DNA → RNA: what do students think the arrow means . CBE Life Sci Educ 13 , 338-348. Link , Google Scholar
- Yoshikawa H, Weisner TS, Kalil A, Way N ( 2008 ). Mixing qualitative and quantitative research in developmental science: uses and methodological choices . Dev Psychol 44 , 344-354. Medline , Google Scholar
- Evaluation of the feasibility of a midwifery educator continuous professional development (CPD) programme in Kenya and Nigeria: a mixed methods study 14 May 2024 | BMC Medical Education, Vol. 24, No. 1
- A tutorial for integrating generative AI in mixed methods data analysis 2 August 2024 | Discover Education, Vol. 3, No. 1
- An exploration of the relationship between active learning and student motivation in STEM: a mixed methods study 1 Sep 2024 | Advances in Physiology Education, Vol. 48, No. 3
- Stop the spread: Empowering students to address misinformation through community‐engaged, interdisciplinary science communication training 16 July 2024 | Journal of Research in Science Teaching, Vol. 32
- If you are learner‐centered and you know it, raise your hand: Perspectives on and implementation of pedagogical changes by science instructors during the COVID ‐19 pandemic 27 February 2024 | Biochemistry and Molecular Biology Education, Vol. 52, No. 4
- The Effectiveness of Flipped Classroom Pedagogy in Promoting learning engagement in Organic Chemistry in Grade-12 Students in the Context of South Africa and Covid-19 23 April 2024 | International Journal of Science Education, Vol. 116
- Kamali Sripathi , and
- Aidan Hoskinson
- Ross Nehm, Monitoring Editor
- Digital Capacity Building in Schools: Strategies, Challenges, and Outcomes 18 December 2023 | Médiations et médiatisations, No. 13
- Uptake of, barriers and enablers to the utilization of postnatal care services in Thyolo, Malawi 19 April 2023 | BMC Pregnancy and Childbirth, Vol. 23, No. 1
- Using mixed methods to understand and tackle barriers to accessing health services 21 Aug 2023 | Revista Panamericana de Salud Pública, Vol. 47
- The Impact of Mobile Learning on Students’ Attitudes towards Learning in an Educational Technology Course 20 July 2023 | Multimodal Technologies and Interaction, Vol. 7, No. 7
- Farmers Perceptions on Artificial Insemination (AI): A Mixed Method Design 8 June 2023 | European Journal of Theoretical and Applied Sciences, Vol. 1, No. 3
- Melissa L. Aikens and
- Alexander R. Kulacki
- Brian Sato, Monitoring Editor
- When the school door closes, do teachers open a window? Using diary method to investigate teachers’ online teaching practices and momentary experiences in crisis 1 Mar 2023 | Computers & Education, Vol. 194
- Strategies to Increase Willingness to Receive a COVID-19 Vaccine among Nursing Home Staff 31 December 2022 | Infectious Disease Reports, Vol. 15, No. 1
- Undergraduate students’ approaches to learning biology: a systematic review of the literature 26 November 2021 | Studies in Science Education, Vol. 59, No. 1
- İlkokul Öğrencilerinin Beslenme Çantası İçeriklerinin Farklı Değişkenler Açısından İncelenmesi 30 December 2022 | Gazi Üniversitesi Gazi Eğitim Fakültesi Dergisi, Vol. 42, No. 3
- A case study of a novel summer bridge program to prepare transfer students for research in biological sciences 20 December 2022 | Disciplinary and Interdisciplinary Science Education Research, Vol. 4, No. 1
- Dance in Zoom: Using video conferencing tools to develop students' 4C skills and self-efficacy during COVID-19 1 Dec 2022 | Thinking Skills and Creativity, Vol. 46
- Using Critical Integrative Argumentation to Assess Socioscientific Argumentation across Decision-Making Contexts 23 September 2022 | Education Sciences, Vol. 12, No. 10
- Erin Stanfield ,
- Corin D. Slown ,
- Quentin Sedlacek , and
- Suzanne E. Worcester
- James Hewlett, Monitoring Editor
- Medical Students’ Perceptions towards Digitization and Artificial Intelligence: A Mixed-Methods Study 13 April 2022 | Healthcare, Vol. 10, No. 4
- Attitudes and preferences of traditional farmers toward reproductive biotechnology application for improved indigenous pig production in Zambia 22 February 2022 | Veterinary World
- PhD experience (and progress) is more than work: life-work relations and reducing exhaustion (and cynicism) 25 March 2020 | Studies in Higher Education, Vol. 47, No. 2
- Mixed Methods and Quality of Postgraduate Research 1 Jan 2022
- The effect of a smartphone-based pain management application on pain intensity and quality of life in adolescents with chronic pain 23 March 2021 | Scientific Reports, Vol. 11, No. 1
- Paired Multiple-Choice Questions Reveal Students’ Incomplete Statistical Thinking about Variation during Data Analysis 10 Sep 2021 | Journal of Microbiology & Biology Education, Vol. 22, No. 2
- An Interprofessional E-Learning Resource to Prepare Students for Clinical Practice in the Operating Room—A Mixed Method Study from the Students’ Perspective 11 August 2021 | Healthcare, Vol. 9, No. 8
- Towards an Open Medical School without Checkerboards during the COVID-19 Pandemic: How to Flexibly Self-Manage General Surgery Practices in Hospitals? 17 June 2021 | Healthcare, Vol. 9, No. 6
- Building Excellence in Scientific Teaching: How Important Is the Evidence for Evidence-Based Teaching when Training STEM TAs? 30 Apr 2021 | Journal of Microbiology & Biology Education, Vol. 22, No. 1
- Growing a growth mindset: characterizing how and why undergraduate students’ mindsets change 8 July 2020 | International Journal of STEM Education, Vol. 7, No. 1
- Scientist Spotlights: Online assignments to promote inclusion in Ecology and Evolution 8 October 2020 | Ecology and Evolution, Vol. 10, No. 22
- Emily E. Scott ,
- Mary Pat Wenderoth , and
- Jennifer H. Doherty
- Kimberly Tanner,, Monitoring Editor
- Using concept maps to characterise cellular respiration knowledge in undergraduate students 8 November 2018 | Journal of Biological Education, Vol. 54, No. 1
- Resource Orchestration 1 Jan 2020
- Beyond linear regression: A reference for analyzing common data types in discipline based education research 3 July 2019 | Physical Review Physics Education Research, Vol. 15, No. 2
- Julie Dangremond Stanton ,
- Kathryn Morris Dye , and
- Me’Shae Johnson
- Elisabeth Schussler, Monitoring Editor
- Context-based learning for Inhibition of alternative conceptions: the next step forward in science education 22 June 2018 | npj Science of Learning, Vol. 3, No. 1
- Teachers’ Sensitivity towards Technology and Environmental Ethics 15 August 2018 | Education Sciences, Vol. 8, No. 3
- Simulated Computational Model Lesson Improves Foundational Systems Thinking Skills and Conceptual Knowledge in Biology Students 20 June 2018 | BioScience, Vol. 68, No. 8
- Erin D. Solomon ,
- Michelle D. Repice ,
- Jacinta M. Mutambuki ,
- Denise A. Leonard ,
- Cheryl A. Cohen ,
- Jia Luo , and
- Regina F. Frey
- Cynthia Brame, Monitoring Editor
- How Undergraduate Science Students Use Learning Objectives to Study 1 Jan 2018 | Journal of Microbiology & Biology Education, Vol. 19, No. 2
- Cheryl A. Sensibaugh ,
- Nathaniel J. Madrid ,
- Hye-Jeong Choi ,
- William L. Anderson , and
- Marcy P. Osgood
- Jennifer Momsen, Monitoring Editor
Submitted: 13 January 2016 Revised: 12 June 2016 Accepted: 14 June 2016
© 2016 L. A.-R. M. Warfa. CBE—Life Sciences Education © 2016 The American Society for Cell Biology. This article is distributed by The American Society for Cell Biology under license from the author(s). It is available to the public under an Attribution–Noncommercial–Share Alike 3.0 Unported Creative Commons License (http://creativecommons.org/licenses/by-nc-sa/3.0).

- Science Notes Posts
- Contact Science Notes
- Todd Helmenstine Biography
- Anne Helmenstine Biography
- Free Printable Periodic Tables (PDF and PNG)
- Periodic Table Wallpapers
- Interactive Periodic Table
- Periodic Table Posters
- Science Experiments for Kids
- How to Grow Crystals
- Chemistry Projects
- Fire and Flames Projects
- Holiday Science
- Chemistry Problems With Answers
- Physics Problems
- Unit Conversion Example Problems
- Chemistry Worksheets
- Biology Worksheets
- Periodic Table Worksheets
- Physical Science Worksheets
- Science Lab Worksheets
- My Amazon Books
Qualitative and Quantitative Data – Definitions and Examples

There are two types of data. Qualitative data is descriptive information about characteristics that are difficult to define or measure or cannot be expressed numerically. Quantitative data is numerical information that can be measured or counted.
Qualitative → Qualities
Quantitative → Quantities
Examples of Qualitative Data
- feelings and emotions
- color (unless it can be written as a specific wavelength of light)
- expressions of more/less, ugly/beautiful, fat/thin, healthy/sickly
Examples of Quantitative Data
Quantitative data can be expressed as numbers. If you can measure it, it can be expressed as a quantity.
- number of objects
- temperature
- percentages
Using Both Types of Data
While quantitative data is easier to analyze, qualitative data is also important.
If you are conducting an experiment using the scientific method, for example, you want to record your observations and data as thoroughly as possible.
Let’s say you want to describe a cat. You would use both types of data. Qualitative data could include its fur color and the color of its eyes, whether its fur is soft or rough, the sounds it makes, how it smells, whether it seems affectionate or aloof, its health, whether it is young or old. Quantitative data could include its length, weight, number of toes on each paw, how high it can jump, how many ounces of food it eats each data, and its body temperature.
Related Posts
- Search Menu
Sign in through your institution
- Browse content in Arts and Humanities
- Browse content in Archaeology
- Anglo-Saxon and Medieval Archaeology
- Archaeological Methodology and Techniques
- Archaeology by Region
- Archaeology of Religion
- Archaeology of Trade and Exchange
- Biblical Archaeology
- Contemporary and Public Archaeology
- Environmental Archaeology
- Historical Archaeology
- History and Theory of Archaeology
- Industrial Archaeology
- Landscape Archaeology
- Mortuary Archaeology
- Prehistoric Archaeology
- Underwater Archaeology
- Zooarchaeology
- Browse content in Architecture
- Architectural Structure and Design
- History of Architecture
- Residential and Domestic Buildings
- Theory of Architecture
- Browse content in Art
- Art Subjects and Themes
- History of Art
- Industrial and Commercial Art
- Theory of Art
- Biographical Studies
- Byzantine Studies
- Browse content in Classical Studies
- Classical History
- Classical Philosophy
- Classical Mythology
- Classical Numismatics
- Classical Literature
- Classical Reception
- Classical Art and Architecture
- Classical Oratory and Rhetoric
- Greek and Roman Papyrology
- Greek and Roman Epigraphy
- Greek and Roman Law
- Greek and Roman Archaeology
- Late Antiquity
- Religion in the Ancient World
- Social History
- Digital Humanities
- Browse content in History
- Colonialism and Imperialism
- Diplomatic History
- Environmental History
- Genealogy, Heraldry, Names, and Honours
- Genocide and Ethnic Cleansing
- Historical Geography
- History by Period
- History of Emotions
- History of Agriculture
- History of Education
- History of Gender and Sexuality
- Industrial History
- Intellectual History
- International History
- Labour History
- Legal and Constitutional History
- Local and Family History
- Maritime History
- Military History
- National Liberation and Post-Colonialism
- Oral History
- Political History
- Public History
- Regional and National History
- Revolutions and Rebellions
- Slavery and Abolition of Slavery
- Social and Cultural History
- Theory, Methods, and Historiography
- Urban History
- World History
- Browse content in Language Teaching and Learning
- Language Learning (Specific Skills)
- Language Teaching Theory and Methods
- Browse content in Linguistics
- Applied Linguistics
- Cognitive Linguistics
- Computational Linguistics
- Forensic Linguistics
- Grammar, Syntax and Morphology
- Historical and Diachronic Linguistics
- History of English
- Language Evolution
- Language Reference
- Language Acquisition
- Language Variation
- Language Families
- Lexicography
- Linguistic Anthropology
- Linguistic Theories
- Linguistic Typology
- Phonetics and Phonology
- Psycholinguistics
- Sociolinguistics
- Translation and Interpretation
- Writing Systems
- Browse content in Literature
- Bibliography
- Children's Literature Studies
- Literary Studies (Romanticism)
- Literary Studies (American)
- Literary Studies (Asian)
- Literary Studies (European)
- Literary Studies (Eco-criticism)
- Literary Studies (Modernism)
- Literary Studies - World
- Literary Studies (1500 to 1800)
- Literary Studies (19th Century)
- Literary Studies (20th Century onwards)
- Literary Studies (African American Literature)
- Literary Studies (British and Irish)
- Literary Studies (Early and Medieval)
- Literary Studies (Fiction, Novelists, and Prose Writers)
- Literary Studies (Gender Studies)
- Literary Studies (Graphic Novels)
- Literary Studies (History of the Book)
- Literary Studies (Plays and Playwrights)
- Literary Studies (Poetry and Poets)
- Literary Studies (Postcolonial Literature)
- Literary Studies (Queer Studies)
- Literary Studies (Science Fiction)
- Literary Studies (Travel Literature)
- Literary Studies (War Literature)
- Literary Studies (Women's Writing)
- Literary Theory and Cultural Studies
- Mythology and Folklore
- Shakespeare Studies and Criticism
- Browse content in Media Studies
- Browse content in Music
- Applied Music
- Dance and Music
- Ethics in Music
- Ethnomusicology
- Gender and Sexuality in Music
- Medicine and Music
- Music Cultures
- Music and Media
- Music and Religion
- Music and Culture
- Music Education and Pedagogy
- Music Theory and Analysis
- Musical Scores, Lyrics, and Libretti
- Musical Structures, Styles, and Techniques
- Musicology and Music History
- Performance Practice and Studies
- Race and Ethnicity in Music
- Sound Studies
- Browse content in Performing Arts
- Browse content in Philosophy
- Aesthetics and Philosophy of Art
- Epistemology
- Feminist Philosophy
- History of Western Philosophy
- Metaphysics
- Moral Philosophy
- Non-Western Philosophy
- Philosophy of Language
- Philosophy of Mind
- Philosophy of Perception
- Philosophy of Science
- Philosophy of Action
- Philosophy of Law
- Philosophy of Religion
- Philosophy of Mathematics and Logic
- Practical Ethics
- Social and Political Philosophy
- Browse content in Religion
- Biblical Studies
- Christianity
- East Asian Religions
- History of Religion
- Judaism and Jewish Studies
- Qumran Studies
- Religion and Education
- Religion and Health
- Religion and Politics
- Religion and Science
- Religion and Law
- Religion and Art, Literature, and Music
- Religious Studies
- Browse content in Society and Culture
- Cookery, Food, and Drink
- Cultural Studies
- Customs and Traditions
- Ethical Issues and Debates
- Hobbies, Games, Arts and Crafts
- Natural world, Country Life, and Pets
- Popular Beliefs and Controversial Knowledge
- Sports and Outdoor Recreation
- Technology and Society
- Travel and Holiday
- Visual Culture
- Browse content in Law
- Arbitration
- Browse content in Company and Commercial Law
- Commercial Law
- Company Law
- Browse content in Comparative Law
- Systems of Law
- Competition Law
- Browse content in Constitutional and Administrative Law
- Government Powers
- Judicial Review
- Local Government Law
- Military and Defence Law
- Parliamentary and Legislative Practice
- Construction Law
- Contract Law
- Browse content in Criminal Law
- Criminal Procedure
- Criminal Evidence Law
- Sentencing and Punishment
- Employment and Labour Law
- Environment and Energy Law
- Browse content in Financial Law
- Banking Law
- Insolvency Law
- History of Law
- Human Rights and Immigration
- Intellectual Property Law
- Browse content in International Law
- Private International Law and Conflict of Laws
- Public International Law
- IT and Communications Law
- Jurisprudence and Philosophy of Law
- Law and Politics
- Law and Society
- Browse content in Legal System and Practice
- Courts and Procedure
- Legal Skills and Practice
- Legal System - Costs and Funding
- Primary Sources of Law
- Regulation of Legal Profession
- Medical and Healthcare Law
- Browse content in Policing
- Criminal Investigation and Detection
- Police and Security Services
- Police Procedure and Law
- Police Regional Planning
- Browse content in Property Law
- Personal Property Law
- Restitution
- Study and Revision
- Terrorism and National Security Law
- Browse content in Trusts Law
- Wills and Probate or Succession
- Browse content in Medicine and Health
- Browse content in Allied Health Professions
- Arts Therapies
- Clinical Science
- Dietetics and Nutrition
- Occupational Therapy
- Operating Department Practice
- Physiotherapy
- Radiography
- Speech and Language Therapy
- Browse content in Anaesthetics
- General Anaesthesia
- Clinical Neuroscience
- Browse content in Clinical Medicine
- Acute Medicine
- Cardiovascular Medicine
- Clinical Genetics
- Clinical Pharmacology and Therapeutics
- Dermatology
- Endocrinology and Diabetes
- Gastroenterology
- Genito-urinary Medicine
- Geriatric Medicine
- Infectious Diseases
- Medical Toxicology
- Medical Oncology
- Pain Medicine
- Palliative Medicine
- Rehabilitation Medicine
- Respiratory Medicine and Pulmonology
- Rheumatology
- Sleep Medicine
- Sports and Exercise Medicine
- Community Medical Services
- Critical Care
- Emergency Medicine
- Forensic Medicine
- Haematology
- History of Medicine
- Browse content in Medical Skills
- Clinical Skills
- Communication Skills
- Nursing Skills
- Surgical Skills
- Browse content in Medical Dentistry
- Oral and Maxillofacial Surgery
- Paediatric Dentistry
- Restorative Dentistry and Orthodontics
- Surgical Dentistry
- Medical Ethics
- Medical Statistics and Methodology
- Browse content in Neurology
- Clinical Neurophysiology
- Neuropathology
- Nursing Studies
- Browse content in Obstetrics and Gynaecology
- Gynaecology
- Occupational Medicine
- Ophthalmology
- Otolaryngology (ENT)
- Browse content in Paediatrics
- Neonatology
- Browse content in Pathology
- Chemical Pathology
- Clinical Cytogenetics and Molecular Genetics
- Histopathology
- Medical Microbiology and Virology
- Patient Education and Information
- Browse content in Pharmacology
- Psychopharmacology
- Browse content in Popular Health
- Caring for Others
- Complementary and Alternative Medicine
- Self-help and Personal Development
- Browse content in Preclinical Medicine
- Cell Biology
- Molecular Biology and Genetics
- Reproduction, Growth and Development
- Primary Care
- Professional Development in Medicine
- Browse content in Psychiatry
- Addiction Medicine
- Child and Adolescent Psychiatry
- Forensic Psychiatry
- Learning Disabilities
- Old Age Psychiatry
- Psychotherapy
- Browse content in Public Health and Epidemiology
- Epidemiology
- Public Health
- Browse content in Radiology
- Clinical Radiology
- Interventional Radiology
- Nuclear Medicine
- Radiation Oncology
- Reproductive Medicine
- Browse content in Surgery
- Cardiothoracic Surgery
- Gastro-intestinal and Colorectal Surgery
- General Surgery
- Neurosurgery
- Paediatric Surgery
- Peri-operative Care
- Plastic and Reconstructive Surgery
- Surgical Oncology
- Transplant Surgery
- Trauma and Orthopaedic Surgery
- Vascular Surgery
- Browse content in Science and Mathematics
- Browse content in Biological Sciences
- Aquatic Biology
- Biochemistry
- Bioinformatics and Computational Biology
- Developmental Biology
- Ecology and Conservation
- Evolutionary Biology
- Genetics and Genomics
- Microbiology
- Molecular and Cell Biology
- Natural History
- Plant Sciences and Forestry
- Research Methods in Life Sciences
- Structural Biology
- Systems Biology
- Zoology and Animal Sciences
- Browse content in Chemistry
- Analytical Chemistry
- Computational Chemistry
- Crystallography
- Environmental Chemistry
- Industrial Chemistry
- Inorganic Chemistry
- Materials Chemistry
- Medicinal Chemistry
- Mineralogy and Gems
- Organic Chemistry
- Physical Chemistry
- Polymer Chemistry
- Study and Communication Skills in Chemistry
- Theoretical Chemistry
- Browse content in Computer Science
- Artificial Intelligence
- Computer Architecture and Logic Design
- Game Studies
- Human-Computer Interaction
- Mathematical Theory of Computation
- Programming Languages
- Software Engineering
- Systems Analysis and Design
- Virtual Reality
- Browse content in Computing
- Business Applications
- Computer Security
- Computer Games
- Computer Networking and Communications
- Digital Lifestyle
- Graphical and Digital Media Applications
- Operating Systems
- Browse content in Earth Sciences and Geography
- Atmospheric Sciences
- Environmental Geography
- Geology and the Lithosphere
- Maps and Map-making
- Meteorology and Climatology
- Oceanography and Hydrology
- Palaeontology
- Physical Geography and Topography
- Regional Geography
- Soil Science
- Urban Geography
- Browse content in Engineering and Technology
- Agriculture and Farming
- Biological Engineering
- Civil Engineering, Surveying, and Building
- Electronics and Communications Engineering
- Energy Technology
- Engineering (General)
- Environmental Science, Engineering, and Technology
- History of Engineering and Technology
- Mechanical Engineering and Materials
- Technology of Industrial Chemistry
- Transport Technology and Trades
- Browse content in Environmental Science
- Applied Ecology (Environmental Science)
- Conservation of the Environment (Environmental Science)
- Environmental Sustainability
- Environmentalist Thought and Ideology (Environmental Science)
- Management of Land and Natural Resources (Environmental Science)
- Natural Disasters (Environmental Science)
- Nuclear Issues (Environmental Science)
- Pollution and Threats to the Environment (Environmental Science)
- Social Impact of Environmental Issues (Environmental Science)
- History of Science and Technology
- Browse content in Materials Science
- Ceramics and Glasses
- Composite Materials
- Metals, Alloying, and Corrosion
- Nanotechnology
- Browse content in Mathematics
- Applied Mathematics
- Biomathematics and Statistics
- History of Mathematics
- Mathematical Education
- Mathematical Finance
- Mathematical Analysis
- Numerical and Computational Mathematics
- Probability and Statistics
- Pure Mathematics
- Browse content in Neuroscience
- Cognition and Behavioural Neuroscience
- Development of the Nervous System
- Disorders of the Nervous System
- History of Neuroscience
- Invertebrate Neurobiology
- Molecular and Cellular Systems
- Neuroendocrinology and Autonomic Nervous System
- Neuroscientific Techniques
- Sensory and Motor Systems
- Browse content in Physics
- Astronomy and Astrophysics
- Atomic, Molecular, and Optical Physics
- Biological and Medical Physics
- Classical Mechanics
- Computational Physics
- Condensed Matter Physics
- Electromagnetism, Optics, and Acoustics
- History of Physics
- Mathematical and Statistical Physics
- Measurement Science
- Nuclear Physics
- Particles and Fields
- Plasma Physics
- Quantum Physics
- Relativity and Gravitation
- Semiconductor and Mesoscopic Physics
- Browse content in Psychology
- Affective Sciences
- Clinical Psychology
- Cognitive Psychology
- Cognitive Neuroscience
- Criminal and Forensic Psychology
- Developmental Psychology
- Educational Psychology
- Evolutionary Psychology
- Health Psychology
- History and Systems in Psychology
- Music Psychology
- Neuropsychology
- Organizational Psychology
- Psychological Assessment and Testing
- Psychology of Human-Technology Interaction
- Psychology Professional Development and Training
- Research Methods in Psychology
- Social Psychology
- Browse content in Social Sciences
- Browse content in Anthropology
- Anthropology of Religion
- Human Evolution
- Medical Anthropology
- Physical Anthropology
- Regional Anthropology
- Social and Cultural Anthropology
- Theory and Practice of Anthropology
- Browse content in Business and Management
- Business Ethics
- Business Strategy
- Business History
- Business and Technology
- Business and Government
- Business and the Environment
- Comparative Management
- Corporate Governance
- Corporate Social Responsibility
- Entrepreneurship
- Health Management
- Human Resource Management
- Industrial and Employment Relations
- Industry Studies
- Information and Communication Technologies
- International Business
- Knowledge Management
- Management and Management Techniques
- Operations Management
- Organizational Theory and Behaviour
- Pensions and Pension Management
- Public and Nonprofit Management
- Social Issues in Business and Management
- Strategic Management
- Supply Chain Management
- Browse content in Criminology and Criminal Justice
- Criminal Justice
- Criminology
- Forms of Crime
- International and Comparative Criminology
- Youth Violence and Juvenile Justice
- Development Studies
- Browse content in Economics
- Agricultural, Environmental, and Natural Resource Economics
- Asian Economics
- Behavioural Finance
- Behavioural Economics and Neuroeconomics
- Econometrics and Mathematical Economics
- Economic History
- Economic Systems
- Economic Methodology
- Economic Development and Growth
- Financial Markets
- Financial Institutions and Services
- General Economics and Teaching
- Health, Education, and Welfare
- History of Economic Thought
- International Economics
- Labour and Demographic Economics
- Law and Economics
- Macroeconomics and Monetary Economics
- Microeconomics
- Public Economics
- Urban, Rural, and Regional Economics
- Welfare Economics
- Browse content in Education
- Adult Education and Continuous Learning
- Care and Counselling of Students
- Early Childhood and Elementary Education
- Educational Equipment and Technology
- Educational Strategies and Policy
- Higher and Further Education
- Organization and Management of Education
- Philosophy and Theory of Education
- Schools Studies
- Secondary Education
- Teaching of a Specific Subject
- Teaching of Specific Groups and Special Educational Needs
- Teaching Skills and Techniques
- Browse content in Environment
- Applied Ecology (Social Science)
- Climate Change
- Conservation of the Environment (Social Science)
- Environmentalist Thought and Ideology (Social Science)
- Management of Land and Natural Resources (Social Science)
- Natural Disasters (Environment)
- Pollution and Threats to the Environment (Social Science)
- Social Impact of Environmental Issues (Social Science)
- Sustainability
- Browse content in Human Geography
- Cultural Geography
- Economic Geography
- Political Geography
- Browse content in Interdisciplinary Studies
- Communication Studies
- Museums, Libraries, and Information Sciences
- Browse content in Politics
- African Politics
- Asian Politics
- Chinese Politics
- Comparative Politics
- Conflict Politics
- Elections and Electoral Studies
- Environmental Politics
- Ethnic Politics
- European Union
- Foreign Policy
- Gender and Politics
- Human Rights and Politics
- Indian Politics
- International Relations
- International Organization (Politics)
- Irish Politics
- Latin American Politics
- Middle Eastern Politics
- Political Behaviour
- Political Economy
- Political Institutions
- Political Methodology
- Political Communication
- Political Philosophy
- Political Sociology
- Political Theory
- Politics and Law
- Politics of Development
- Public Policy
- Public Administration
- Qualitative Political Methodology
- Quantitative Political Methodology
- Regional Political Studies
- Russian Politics
- Security Studies
- State and Local Government
- UK Politics
- US Politics
- Browse content in Regional and Area Studies
- African Studies
- Asian Studies
- East Asian Studies
- Japanese Studies
- Latin American Studies
- Middle Eastern Studies
- Native American Studies
- Scottish Studies
- Browse content in Research and Information
- Research Methods
- Browse content in Social Work
- Addictions and Substance Misuse
- Adoption and Fostering
- Care of the Elderly
- Child and Adolescent Social Work
- Couple and Family Social Work
- Direct Practice and Clinical Social Work
- Emergency Services
- Human Behaviour and the Social Environment
- International and Global Issues in Social Work
- Mental and Behavioural Health
- Social Justice and Human Rights
- Social Policy and Advocacy
- Social Work and Crime and Justice
- Social Work Macro Practice
- Social Work Practice Settings
- Social Work Research and Evidence-based Practice
- Welfare and Benefit Systems
- Browse content in Sociology
- Childhood Studies
- Community Development
- Comparative and Historical Sociology
- Disability Studies
- Economic Sociology
- Gender and Sexuality
- Gerontology and Ageing
- Health, Illness, and Medicine
- Marriage and the Family
- Migration Studies
- Occupations, Professions, and Work
- Organizations
- Population and Demography
- Race and Ethnicity
- Social Theory
- Social Movements and Social Change
- Social Research and Statistics
- Social Stratification, Inequality, and Mobility
- Sociology of Religion
- Sociology of Education
- Sport and Leisure
- Urban and Rural Studies
- Browse content in Warfare and Defence
- Defence Strategy, Planning, and Research
- Land Forces and Warfare
- Military Administration
- Military Life and Institutions
- Naval Forces and Warfare
- Other Warfare and Defence Issues
- Peace Studies and Conflict Resolution
- Weapons and Equipment

- < Previous chapter
- Next chapter >
1 What Is Qualitative Research?
- Published: April 2024
- Cite Icon Cite
- Permissions Icon Permissions
Qualitative research methods treat non-numeric empirical materials as data. In other words, Qualitative researchers use words, images, sounds, video, and other data that have not been reduced to numbers to understand some research question of interest. This chapter explores what qualitative research entails and why researchers might want to employ its methods. It begins with a brief history of the emergence of qualitative inquiry. Next, the chapter addresses some of the common misconceptions about qualitative research. It then transitions to an overview of ontology (what is reality) and epistemology (how a person knows a thing) and discusses why understanding one’s own ontological and epistemological preferences is important when approaching how one conducts research. The chapter reviews the use of theory, which is followed by a discussion of decolonizing research.
Personal account
- Sign in with email/username & password
- Get email alerts
- Save searches
- Purchase content
- Activate your purchase/trial code
- Add your ORCID iD
Institutional access
Sign in with a library card.
- Sign in with username/password
- Recommend to your librarian
- Institutional account management
- Get help with access
Access to content on Oxford Academic is often provided through institutional subscriptions and purchases. If you are a member of an institution with an active account, you may be able to access content in one of the following ways:
IP based access
Typically, access is provided across an institutional network to a range of IP addresses. This authentication occurs automatically, and it is not possible to sign out of an IP authenticated account.
Choose this option to get remote access when outside your institution. Shibboleth/Open Athens technology is used to provide single sign-on between your institution’s website and Oxford Academic.
- Click Sign in through your institution.
- Select your institution from the list provided, which will take you to your institution's website to sign in.
- When on the institution site, please use the credentials provided by your institution. Do not use an Oxford Academic personal account.
- Following successful sign in, you will be returned to Oxford Academic.
If your institution is not listed or you cannot sign in to your institution’s website, please contact your librarian or administrator.
Enter your library card number to sign in. If you cannot sign in, please contact your librarian.
Society Members
Society member access to a journal is achieved in one of the following ways:
Sign in through society site
Many societies offer single sign-on between the society website and Oxford Academic. If you see ‘Sign in through society site’ in the sign in pane within a journal:
- Click Sign in through society site.
- When on the society site, please use the credentials provided by that society. Do not use an Oxford Academic personal account.
If you do not have a society account or have forgotten your username or password, please contact your society.
Sign in using a personal account
Some societies use Oxford Academic personal accounts to provide access to their members. See below.
A personal account can be used to get email alerts, save searches, purchase content, and activate subscriptions.
Some societies use Oxford Academic personal accounts to provide access to their members.
Viewing your signed in accounts
Click the account icon in the top right to:
- View your signed in personal account and access account management features.
- View the institutional accounts that are providing access.
Signed in but can't access content
Oxford Academic is home to a wide variety of products. The institutional subscription may not cover the content that you are trying to access. If you believe you should have access to that content, please contact your librarian.
For librarians and administrators, your personal account also provides access to institutional account management. Here you will find options to view and activate subscriptions, manage institutional settings and access options, access usage statistics, and more.
Our books are available by subscription or purchase to libraries and institutions.
| Month: | Total Views: |
|---|---|
| May 2024 | 5 |
| June 2024 | 5 |
| July 2024 | 8 |
| August 2024 | 5 |
- About Oxford Academic
- Publish journals with us
- University press partners
- What we publish
- New features
- Open access
- Rights and permissions
- Accessibility
- Advertising
- Media enquiries
- Oxford University Press
- Oxford Languages
- University of Oxford
Oxford University Press is a department of the University of Oxford. It furthers the University's objective of excellence in research, scholarship, and education by publishing worldwide
- Copyright © 2024 Oxford University Press
- Cookie settings
- Cookie policy
- Privacy policy
- Legal notice
This Feature Is Available To Subscribers Only
Sign In or Create an Account
This PDF is available to Subscribers Only
For full access to this pdf, sign in to an existing account, or purchase an annual subscription.
Loading metrics
Open Access
Peer-reviewed
Research Article
Model-Based Analysis for Qualitative Data: An Application in Drosophila Germline Stem Cell Regulation
Affiliation Weldon School of Biomedical Engineering, Purdue University, West Lafayette, Indiana, United States of America
Affiliation Department of Mathematics, Purdue University, West Lafayette, Indiana, United States of America
* E-mail: [email protected]
Affiliation Agricultural and Biological Engineering, Purdue University, West Lafayette, Indiana, United States of America
- Michael Pargett,
- Ann E. Rundell,
- Gregery T. Buzzard,
- David M. Umulis
- Published: March 13, 2014
- https://doi.org/10.1371/journal.pcbi.1003498
- Reader Comments
Discovery in developmental biology is often driven by intuition that relies on the integration of multiple types of data such as fluorescent images, phenotypes, and the outcomes of biochemical assays. Mathematical modeling helps elucidate the biological mechanisms at play as the networks become increasingly large and complex. However, the available data is frequently under-utilized due to incompatibility with quantitative model tuning techniques. This is the case for stem cell regulation mechanisms explored in the Drosophila germarium through fluorescent immunohistochemistry. To enable better integration of biological data with modeling in this and similar situations, we have developed a general parameter estimation process to quantitatively optimize models with qualitative data. The process employs a modified version of the Optimal Scaling method from social and behavioral sciences, and multi-objective optimization to evaluate the trade-off between fitting different datasets (e.g. wild type vs. mutant). Using only published imaging data in the germarium, we first evaluated support for a published intracellular regulatory network by considering alternative connections of the same regulatory players. Simply screening networks against wild type data identified hundreds of feasible alternatives. Of these, five parsimonious variants were found and compared by multi-objective analysis including mutant data and dynamic constraints. With these data, the current model is supported over the alternatives, but support for a biochemically observed feedback element is weak (i.e. these data do not measure the feedback effect well). When also comparing new hypothetical models, the available data do not discriminate. To begin addressing the limitations in data, we performed a model-based experiment design and provide recommendations for experiments to refine model parameters and discriminate increasingly complex hypotheses.
Author Summary
We developed a process to quantitatively fit mathematical models using qualitative data, and applied it in the study of how stem cells are regulated in the fruit fly ovary. The available published data we collected are fluorescent images of protein and mRNA expression from genetic experiments. Despite lacking quantitative data, the new process makes available quantitative model analysis techniques to reliably compare different models and guide future experiments. We found that the current consensus regulatory model is supported, but that the data are indeed insufficient to address more complex hypotheses. With the quantitatively fit models, we evaluated hypothetical experiments and estimated which future measurements should best refine or test models. The model fitting process we have developed is applicable to many biological studies where qualitative data are common, and can accelerate progress through more efficient experimentation.
Citation: Pargett M, Rundell AE, Buzzard GT, Umulis DM (2014) Model-Based Analysis for Qualitative Data: An Application in Drosophila Germline Stem Cell Regulation. PLoS Comput Biol 10(3): e1003498. https://doi.org/10.1371/journal.pcbi.1003498
Editor: Jeffrey J. Saucerman, University of Virginia, United States of America
Received: August 20, 2013; Accepted: January 16, 2014; Published: March 13, 2014
Copyright: © 2014 Pargett et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License , which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Funding: This research was sponsored in part by the National Science Foundation (NSF, http://www.nsf.gov/ ) CAREER Award to AER and funded under the American Recovery and Reinvestment Act of 2009 (ARRA) under the grant ECCS-0846572 and a grant from the National Institute of Health (NIH: HD73156) to DMU. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Competing interests: The authors have declared that no competing interests exist.
Introduction
Biological systems are often characterized using qualitative data, such as stained images, immunoblots, microarrays, or observations of cell morphology, rather than absolute values (e.g. molecular concentration). Such qualitative data typically show relative relationships in how a system characteristic (e.g. expression of proteins or mRNA, morphology, phenotype) is distributed spatially, and/or changes with time or with genetic perturbations. These data are prevalent due to the complexity of biological systems and measurements, from spatial organization and dynamic behavior, to the need for multi-step reactions to generate a measurable signal, along with the wide variability of experimental factors (e.g. reagent concentrations, background interference, antibody quality and specificity) [1] , [2] . In many cases, more time-consuming quantifiable measurements are sacrificed for improved throughput and spatial resolution [3] , though the resulting uncertainty in absolute value, range and resolution is limiting, particularly as applied in mathematical models. In this study, we address these limitations in model-data integration in the context of a stem cell niche in the Drosophila germarium, as the available data are largely qualitative and it has become a model system from which we hope to gain insight into stem cell regulation.
Illustrated in Figure 1A , each oblong germarium houses 2–3 germline stem cells (GSC) associated with the cap cells (CC) at the anterior end. In the course of differentiating, GSC progeny transition through a cystoblast phase (CB, single cells beginning to express differentiation factors), then divide repeatedly forming cysts interconnected by a fusome. In undifferentiated cells, the fusome structure is isolated and spherical, referred to as a spectrosome (fusome/spectrosome morphology is a common observation). The regulation of stem cell self-renewal vs. differentiation depends on signaling by Decapentaplegic (Dpp), a bone morphogenic protein (BMP) ligand homologue, which is expressed by the cap cells. As illustrated in Figure 1B , the GSC is maintained by Dpp signaling, mediated through surface receptors that promote phosphorylation of Mad to pMad. pMad acts as an input to a regulatory network, including (at least) Bam, Nos and Brat. For more complete coverage of germarium structure and function, we recommend recent reviews [4] – [6] .
- PPT PowerPoint slide
- PNG larger image
- TIFF original image
A) Diagram of Wild Type germarium structure with anterior to the left, showing cap cells (CC), germline stem cells (GSC), cystoblasts (CB), and cysts. Below, a color-matched schematic of the 1 dimensional model used in this study. B) Diagram of signaling in the anterior germarium, showing the internal regulatory state of the GSC (left), and CB (right). Yellow and blue boxes refer to differentiation- and self-renewal-promoting elements, respectively. C) Examples of typical images (upper) and qualitative interpretations (lower) comprising available data. Qualitative interpretations are mappings of relative intensity (fluorescent, colorimetric, etc.) and original author interpretations to the 1-D model, indicated at bottom. Relevant color channels per image are (from left to right): pMad in red, Nos in red, Bam in green, Phenotype showing spectrosomes in red. Images reproduced/adapted from [57] (1st from left), [58] (2nd and 3rd from left), and [59] (right).
https://doi.org/10.1371/journal.pcbi.1003498.g001
Multiple types of data inform GSC regulation in the germarium. Most prominently, spatial distribution data are published in the form of fluorescent intensity for several proteins (in some cases mRNA) in wild type and in different mutant backgrounds (examples shown in Figure 1C ). Were all of these data collected via fully quantitative techniques, they would be approachable with common model fitting techniques (e.g. least squared error regression). However, measuring precise quantitative levels of protein or mRNA in vivo continues to be very technically challenging, and the existing qualitative data are not directly comparable for optimizing models by typical regression. The scaling of fluorescent intensity differs for each molecular species, relying on different antibodies and the reaction conditions for each sample, and in some instances data are aggregated from different publications. Individually, these qualitative data provide loose constraints and to be effective they must be considered simultaneously, for which new methods are needed. Furthermore, the data are provided by three principle types of observations: wild type protein distributions at a single time point, distributions for different subsets of the components in mutant and/or ectopic expression experiments, and estimates of the time between cell cycles that provide a dynamic constraint. Lacking further information, it is unclear if one of these disparate observation types is better for a model to satisfy than another, which leaves only limited meaning to a single best parameter set.
To assess feasible mechanisms in this system (and others with qualitative constraints), we developed an integrated strategy comprising two applications seldom used with biological models: Optimal Scaling to quantitatively estimate model fitness, and Pareto multi-objective optimization to simultaneously consider multiple disparate types of data. Though we apply it here in a study of stem cell regulation in the Drosophila germarium, the procedure is applicable to any quantitative model.
This study integrates research in three primary areas: (1) optimization with qualitative data and the Optimal Scaling procedure, (2) the problem of using multiple disparate datasets, and multi-objective optimization as a robust solution, and (3) modeling of stem cell regulation in the Drosophila germarium. The remainder of this introduction is dedicated to informing these three areas.
Optimization and qualitative data: Optimal Scaling
Optimization algorithms attempt to find a parameter set (or point, i.e. a value for each uncertain parameter) that gives the best value for some objective defining model fitness, typically the error between model predictions and data; they are commonly identified as either local or global methods ( Figure 2B illustrates these as applied in this study). Local optimization starts at a specific parameter set and selects a search direction and step based on the gradient, i.e. how much the error changes with small parameter changes. Global methods use the fitness evaluated for a sampling of parameter sets to then select new samples expected to improve (algorithmic details vary). Qualitative data, such as the fluorescent images of the germarium, define predominantly binary fitness criteria; either the model outputs satisfy the observation or not. They provide no gradient information and discontinuous changes in fitness that may be difficult to identify. Optimization procedures are likely to fail to see where a better solution might lie if a sample did not happen to be placed there. As a result, biological model parameters have typically been estimated either using only data that is quantitative, or by the modeler manually adjusting parameters based on intuition, a very time-consuming process.
https://doi.org/10.1371/journal.pcbi.1003498.g002
To design a general procedure for optimization to qualitative data, we considered past efforts in several fields that have addressed aspects of the problem. We predominantly build on the Optimal Scaling method, reviewed below, but it is informative to comment on alternative techniques available. In statistics, regression to qualitative data has a long history [7] , but in contrast with the mechanistic biological context, only minimal models are used. These statistical models are typically linear with some assumed structure on the data (i.e. a function such as logit or probit is applied to the model values). Thresholds are defined to subdivide the continuous model output into intervals, and map each interval to a discrete qualitative output (e.g. high and low, or a phenotype name). The reliance on model linearity limits the immediate utility of past statistical approaches for the non-linear models at hand. In complex model analysis, behavior discrimination [8] has recently been described to define thresholds among different model behaviors, but could be applied to model tuning with qualitative data. It relies on mathematical descriptions of each qualitative behavior to create quantitative metrics to evaluate how near a model is to satisfying each behavior. Defined behaviors can range from simple thresholds to complex time-dependent relationships. A conceptual compromise, Optimal Scaling [9] is an older approach that originated in the social sciences. Similar to behavior discrimination, it evaluates a distance from the point of satisfaction, but is more directly oriented toward model tuning. It also resembles statistical regression problems, but while its past use has been with simple models, it is more generally applicable to complex cases (i.e. non-linear models). For a particular model output, Optimal Scaling uses regression to estimate the optimal quantitative values likely to have generated the qualitative data, i.e. the best-case fit to that model output. While each of these approaches estimates a quantitative fitness, Optimal Scaling offers particularly broad applicability and a focus on the feasible values of the real system.
The Optimal Scaling process is illustrated in Figure 2A , and details are provided in Methods . Each time a model output is considered, Optimal Scaling defines quantitative values to replace the qualitative observations; we refer to these as surrogate data (illustrated as blue circles in Figure 2A , right). The surrogate data are intended to represent what could have existed in the true system. To evaluate the best-case fitness to the given model output, the surrogate data values are optimized within the constraint that they still satisfy the qualitative observations (constraints shown as shaded boxes in Figure 2A ). The quantitative error between these optimal values and the model output then defines the model fitness, and may be used as the objective for existing optimization techniques. As originally presented for regression of simple models in the social sciences, Optimal Scaling is alternated with a least squares optimization of parameter values [9] , [10] . However, for more complex models, the necessary convexity of that optimization scheme can not be guaranteed. Instead, to apply global and multi-objective optimization techniques, we nest the optimal scaling step fully within the parameter estimation problem [1] (i.e. optimal scaling is performed explicitly for every parameter set evaluated). For details on the optimization process, see Methods .
Using disparate datasets: Multi-objective optimization
The Optimal Scaling procedure addresses model fitness to the qualitative distributions from germarium images (examples in Figure 1C ), but the uncertainty among the different observation types remains. For quantitative data, the trade-off between satisfying each type would be informed by measured experimental variance. For these qualitative data, we suggest that the problem can be viewed as having multiple objectives, i.e. fitting each type of data as a separate objective (as described in general in [1] ). In this way, the risk of bias in estimating a single best parameter set is mitigated and a more complete perspective on model performance constructed by evaluating the continuous trade-off among fitting the different data types. An approach that originated in economics and is commonly applied in design optimization, the multi-objective Pareto optimality concept focuses on determining a well-spaced set of points describing this trade-off, each of which corresponds to an optimal point for a different weighting among objectives [11] . Therein, a point is considered Pareto optimal if no other points improve one objective without compromising another.
Evaluating a set of Pareto optimal points (termed the Pareto front, demonstrated in Figure 2C ) comes at a significant computational cost. It is useful to minimize the dimension of the multiobjective problem and group the most similar data together. While we consider the germarium data grouped by the type of observation, data can be grouped in a variety of other ways as suits the problem at hand, including the quality of data (e.g. nominal, ordinal, ratiometric, etc.), or the measurement technique used. The Pareto front is described by plotting the Pareto optimal points on the objective space (e.g. fitness to wild type data vs. to mutant data). Reflected in its placement and curvature, the Pareto front shows the trade-off between objectives, such as how much wild type fitness must be sacrificed to better fit mutants. Accordingly, we can then use the Pareto front to compare the performance of different models.
In this study we analyze simplified spatio-temporal models of the germarium subject to a compiled group of available qualitative data by estimating quantitative fitness through Optimal Scaling. To robustly capture data-consistent model behavior, we use multi-objective optimization to estimate a group of representative model parameter sets (Representatives). With this approach, we are able to refine predictive estimates of system behavior, discriminate among multiple models, and estimate the merit of future experiments. To develop and demonstrate the approach in the germarium, we compare alternative regulatory networks generated by a naive screen, as well as mechanistic hypotheses informed by current evidence, including a model based on previous work [12] . We then estimate Representative parameter sets (Pareto points, in this study) for each model and discriminate among models based on their simultaneous fitness to published qualitative protein and mRNA distribution data from wild type and mutant organisms. Using the Representatives for each model, we assess which data and parameters should be considered in expanding on the current models, and estimate which future experiments will be most informative by model-based experiment design.
Results/Discussion
Stem cell regulation in the germarium has been represented by a variety of conceptual models [4] – [6] along with a spatio-temporal mathematical model [12] . Among these, the consensus regulatory network most widely supported is as shown in Figure 1B , referred to herein as the Core network. pMad, Bam, Nos, and Brat form a chain of repressors that results in bistable behavior with either pMad and Nos present (the self-renewing state, Figure 1B , left) or Bam and Brat present (the differentiating state, Figure 1B , right). This network is well supported through genetic and protein interaction studies, as well as modeling analysis [12] , [13] and serves as the starting point for the mathematical modeling further developed herein. As indicated by question marks in Figure 1B , additional components and interactions are unknown, but expected. Examples include miRNA mediated repression [14] , ligand endocytosis [15] , [16] , modification of the extracellular environment [17] – [20] , and a variety of cell contact mediated mechanisms [21] , [22] .
Compiled protein expression data
Data were compiled from published images of protein expression across wild type and mutant germaria. All of the data used ( Table 1 ) are qualitative, giving relative expression of proteins, as shown in Figure 1 . Phenotype data are common and indicate fusome morphology (e.g. Figure 1C , image on right, showing in red the spectrosomes as round and fusomes as branched). We correlate the fusome development to Brat expression as an indicator of differentiation. We consider the germarium divided into 4 regions: GSC, CB, Cyst and Posterior. The mapping of these regions onto our 1 dimensional models is shown in Figure 1A lower, indicated by color (See also Figure S1 in Supporting Information, Text S1 , and refer to Methods for modeling). We provide this color map as a reference for model outputs throughout the analysis.
https://doi.org/10.1371/journal.pcbi.1003498.t001
Example qualitative interpretations of data are provided below each image in Figure 1C , with a reference to the 1-D model. We note that Bam is known to be repressed by RBP9, which is present in the posterior region of the germarium [23] . However, the regulation of RBP9 remains unknown. We neglect this posterior repression on Bam in the data, as it is outside the scope of the models we test. Examining the qualitative interpretations, it is apparent that each observation provides only loose constraints, emphasizing the importance of considering many such observations simultaneously.
To separate the different types of observations used, we divide data among three categories and independently evaluate model satisfaction of (1) Wild Type observations, (2) Mutant observations, and (3) Behavioral observations. The Behavioral category includes both dynamic constraints, specifying how quickly the cells must respond, and negative phenotypes observed in mutants, which reflect robustness to some perturbations (indicated in Table 1 ). These categories were chosen both for biological interest and to aggregate data expected to be similar. For example, Mutants commonly exhibit an all or nothing response over the entire germarium, while Wild Type responses are more graded.
Representative parameter estimation procedure
We developed a new approach to search for Representatives that best satisfy qualitative data, which incorporates three elements: (1) the novel application of Optimal Scaling to quantitatively estimate model fitness, (2) global optimization to select a single best solution for each objective, and (3) multi-objective optimization to find a set of Representatives irrespective of weighting among objectives. Our implementation of these techniques is illustrated in Figure 2 . For details on each of these processes, consult Methods .
The quantification of model fitness by Optimal Scaling in this study is represented in Figure 2A . The procedure generates surrogate data (blue circles) that are required to lie within intervals that ensure consistency with qualitative data (shaded boxes). Model error is then calculated as a relative sum of squared error between surrogates and the model output (green line). Note that error is only non-zero when surrogates cannot be perfectly aligned with the model output, as in cell positions 3 and 4 in Figure 2 A . The optimization problem in Optimal Scaling is to select the intervals and surrogates that minimize the model error for a given model output.
The global parameter estimation process is depicted in Figure 2B . In this study, we address non-linear spatio-temporal systems with a minimum of 10 states and 18 uncertain parameters. When estimating parameters, dense parameter screening is infeasible and gradient-based searches are not expected to reliably arrive at a global solution, but identify local optima instead. To proceed, we employ a hybrid semi-deterministic approach comprising a sparse global screen followed by a multi-start gradient search. It is important to keep in mind that for these models, available optimization techniques do not guarantee globally optimal or unique solutions (note the unidentified local minimum in Figure 2B right).
Finally, to generate the set of representative model parameters, we use multi-objective optimization to find points on the Pareto front, as illustrated in Figure 2C . Here, we determine the Pareto points (the Representatives) using the Normalized Normal Constraint (NNC) method [24] ( Figure 2C , right), with modifications to suit the problem at hand and take advantage of global screening (see Methods for more details). This method performs multiple single-objective gradient searches, with each restricted to lie on a different line so that resulting points are well spaced (dashed lines in Figure 2C , right).
Germarium Core network performance
The multi-objective approach reliably determines a set of Representatives for the germarium models. The Pareto front identified for the Core regulatory network (as depicted in Figure 1B ) is shown in Figure 3 .
https://doi.org/10.1371/journal.pcbi.1003498.g003
The front is quite convex (toward the Utopian point), but with a significant trade-off between Wild Type and Mutant fitness (2nd from left). Behavioral fitness closely matches Wild Type (3rd from left, note the very small scale) and exhibits a similar trade-off with Mutant fitness (right). To illustrate fitness, Figure 3B presents examples of both well and poorly fit observations, for the Pareto point nearest the Utopian (arrows in Figure 3A ), chosen by Euclidean distance to estimate a midpoint in the trade-off (fitness at nearby Pareto points was similar, data not shown). Most of the observations are satisfied, or nearly satisfied, at this point. The two largest misfits are pMad in a dMyc mutant with ectopic dMyc expression, and pMad in a Brat mutant (arrows in Figure 3B ).
Examining the data and results for the Brat mutant leads to two important comments. First, we note that the interpretation of the Brat mutant phenotype may be overly aggressive (i.e. too many cells designated with high pMad), due to the discretization of the germarium into the 4 regions considered in this study. The Cyst region extends throughout the 2–8 cell cysts (cell 3–9 in the 1D model), but the indications from data of high pMad expression past the CB do not clearly extend throughout 8 cell cysts [12] . Second, while pMad signaling in the Brat mutant extends beyond the CB, that in Bam mutants does not [25] , suggesting either an unknown regulatory interaction or inconsistency among experiments.
Network inference supports the core structure
To evaluate the Core model in our framework, we compare alternative connections of its regulatory elements, pMad, Bam, Nos, and Brat (different model structures, i.e. rewiring of network edges). Through a simple network inference problem focused on Wild Type data only, we performed a broad screen of alternative networks and identified a set of feasible networks to more thoroughly evaluate, shown in Figure 4 . Considering only Wild Type fitness, we tested the ∼65 k alternatives with only inhibitory connections and additionally performed searches for alternatives that include activation, beginning with 250 k samples. Refer to Methods and Supporting Information ( Text S1 ) for details. Due to the sparse qualitative data, many networks (hundreds) were identified as capable of fitting Wild Type data. To refine this large group, we relied on the principle of parsimony, preferring simple networks (i.e. those with fewer connections). Most of the acceptable networks were nested (i.e. contained simpler acceptable networks plus additional connections). From these, we identified five parsimonious variants containing no simpler acceptable networks. We additionally included two networks with extra connections, chosen arbitrarily, to provide a comparison for trade-offs in more complex, but uninformed, models. (‘Alt6’, ‘Alt7’). We compared these networks against the Core, using all available data to generate a Pareto front for each.
Yellow and blue boxes refer to differentiation- and self-renewal-promoting elements, respectively.
https://doi.org/10.1371/journal.pcbi.1003498.g004
Pareto fronts determined for each of the alternative networks are shown superimposed in Figure 5A (between 11 and 46 Representatives per network). All networks fail to fully satisfy the data. Examining the Wild Type vs. Mutant projection to compare performance among networks, the Core network dominates most alternatives ( Figure 5A , left). However, networks Alt1 and Alt4 perform very similarly to the Core, dominating it at some points. To more closely compare these three models, we examine fitness to individual data ( Figure 5B , plots from the Representatives nearest the Utopian, arrows in Figure 5A ). For reference, we also present results from Alt3, which performs poorly (e.g. compare top plots, where Nos is observed uniformly high in Bam mutants). In the Nos mutant where Brat data are uniformly high, Alt3 fails while Alt4 performs quantitatively better ( Figure 5B 2nd row). However, the qualitative decrease in the anterior region for Alt4 indicates that its structure is less consistent with the mutant phenotype than the Core or Alt1 (compare 2nd plots for each network, decrease in Alt4 indicated by arrow).
A) Solved Pareto fronts for naively generated networks, superimposed for comparison. Insets enlarge view near Utopian point. B) Examples of model fitness, comparing the Core network with the two closest alternatives (based on Pareto front placement) and a poor alternative. Only Alt1 remains similar to the Core on examination. Simulated from Pareto points closest to the Utopian, black arrows in A. Notation as in Figures 3 and 2A .
https://doi.org/10.1371/journal.pcbi.1003498.g005
The Core and Alt1 networks perform quite similarly, with only minor differences among the unfit data ( Figure 5B , compare 3rd and bottom plots). All of the networks compared failed to fit these data. The similar performance of these two networks is explained by similarity in structure. The only difference between the two is that Alt1 lacks feedback of Brat upon Mad ( Figure 4A ). The basic structure of the Core network is thus well supported, but the data provide poor support for the feedback component.
These comparisons and the relative lack of support for feedback exemplify how sparse and qualitative data can be limiting, even when evaluated quantitatively. Rather than suggesting that the well observed feedback element is not involved, this study indicates that the readily available data from genetic experiments are not sensitive to feedback on Mad. Instead, biochemical evidence indicates the repression of Mad in the presence of Brat (with Pumilio as a cofactor) in a Drosophila S2 cell line [12] . While such data can be applied directly to define a model, it is not an explicit observation of the germarium that can be compared to simulations. Furthermore, to better understand the system and build parsimonious models, we encourage considering feasible alternatives to the observed interactions, and asking what is necessary for the system to function, i.e. if elements are indispensable, redundant or unimportant. The example experiment design provided below suggests other genetic experiments in the germarium that may be more sensitive to the feedback on Mad.
Data do not discriminate more complex hypothesized networks
We constructed four hypothetical networks that include additional regulatory mechanisms, as discussed in recent literature. Each contains the Core network along with additional components and interactions ( Figure 6A ). For simplicity with the current model structure, we do not consider mechanisms based on cell-cell contact and adhesion.
A) Informed hypothetical networks, each adding elements to the Core network. * indicates an indirect interaction, potentially involving many intermediaries. Coloring as previously, with white boxes where effects are not conceptually clear. B) Solved Pareto fronts for informed hypothetical networks, including Alt1 which performs similarly to the Core. Arrows indicate the only distinguishable feature among them, where Ago1 improves over other networks, though only at one point.
https://doi.org/10.1371/journal.pcbi.1003498.g006
- The endocytosis mediated network Endo introduces feedback on the cellular endocytosis rate, via Brat inhibiting dMyc production, as previously observed [12] . dMyc promotes endocytosis [15] , [16] , which hypothetically causes a stem cell to internalize ligand-receptor complexes more quickly, creating competition for ligand.
- The Piwi mediated network Piwi includes feedback via Piwi and the associated piRNAs, hypothesized to repress Smurf [26] , a ubiquitin protein ligase, which promotes the degradation of both pMad [25] , [27] , [28] and cell-surface receptors for Dpp [29] , [30] (in conjunction with an unmodeled cofactor, Fused). Piwi is repressed in Bam expressing cells [31] , though direct interaction with Bam has not been investigated. Because Piwi is in the Argonaute family [32] , whose members in several organisms associate with proteins bearing TRIM-NHL motifs (e.g. Brat, Mei-P26 in Drosophila and TRIM32 in mice) [14] , [33] , this model places Piwi as a hypothetical target of Brat.
- In the Argonaute mediated network Ago, feedback is hypothetically mediated by the observed Mei-P26 repression of miRNA levels (interacting with Argonaute-1) [14] , taking effect through miR-184, which represses a Dpp receptor as well as pMad signaling [34] . Not all TRIM-NHL protein interactions involve degradation activity [35] , so Argonaute-1 itself may not be directly regulated by Mei-P26. However, for simplicity in this network, Ago represents the overall function of Argonaute-1 and miR-184 and is regulated by Mei-P26.
- The diffusion mediated network Diff regards extracellular modification of effective Dpp diffusivity through its association with both collagen and the proteoglycan Dally [17] – [19] . Germline cells are hypothesized to regulate the expression of Dally in nearby somatic cells through endothelial growth factor (EGF) ligands [20] ; herein, the regulation of EGF is placed downstream of Brat, as the hypothetical regulator of differentiation processes. Dpp association with either collagen or Dally limits its diffusivity, with Dally expression modifying the pool of binding sites to retain Dpp nearby a given germline cell.
We include both the Core and Alt1 networks in the analysis, as they perform nearly indistinguishably. Pareto fronts are presented superimposed in Figure 6B . As indicated by the overlap of all fronts, no clear improvements are made by the hypothetical networks, based on the data at hand. The only indication of improved fitness is a lower error achieved by the Ago model for Wild Type and Behavioral data, while relatively well fit to Mutant data (examine left and right plots, respectively, at the Mutant anchor point indicated by arrows). However, no clear improvements are apparent in individual outputs for the Ago model (data not shown). The lack of clear discrimination among models indicates that the currently available data is inadequate to distinguish the expanded mechanisms tested.
Analysis of hypothetical networks for future development
Beyond model performance, we use the identified Representatives (Pareto points) to assess the relative influence of each observation and parameter as we consider future model development. We examine the distribution of model error to identify which observations are not yet consistently satisfied, and the distribution of parameter sensitivity to identify influential parameters.
Few data remain poorly fit.
Data that are consistently well fit across all models need no further attention (unless quantitative measurements become feasible). A useful alternative perspective is that these data are not capable of distinguishing between current and more complex models. Consistently well fit data include indices 5, 7, 8, 13, 14, and 17 from Table 1 : the phenotype, Nos or Brat in Bam −/−, pMad in dMyc overexpression, Brat in Nos −/−, and the phenotype for Dpp −/−. The few data consistently unfit, which may prove useful in considering future model additions include 11, 15 and 18 in Table 1 : pMad in dMyc −/− with dMyc overexpression, pMad in Brat −/−, and the phenotype in Dpp +/−. The remaining data were involved in the tradeoff between Wild Type and Mutant objectives, and will remain useful when evaluated simultaneously. The error distributions examined are available in Supporting Information (Figures S2–S8 in Text S1 ).
Regulatory parameters exert the greatest effect.
We use local sensitivity analysis to measure the impact of individual model parameters, though it is specific to each Representative. As expected, the half-maximal concentrations characterizing regulatory interactions produce the most significant effects and are heavily involved in the trade-off among data objectives. Also indicated as important are phosphorylation kinetics for Mad, and the diffusion, binding and degradation of Dpp. Degradation of regulators and dissociation of Dpp from receptors rarely have a significant effect. Distributions of parameter sensitivity are available in Supporting Information (Figures S9–S14 in Text S1 ).
Experiment design
Using the Representatives, we are able to perform a simple model based experiment design, aiming to estimate the most informative experiments from a set of hypothetical perturbations and measurements. Each Representative of each model produces an individual estimate of the system response in a novel experiment. Potential experiments can then be selected to reduce uncertainty in model parameters, in model outputs or to discriminate among competing model structures.
To consider different expectations from data as well as different modeling goals, we present a small variety of approaches to the experiment design problem. First, we focus on a realistic case, expecting qualitative protein distributions, as with current data. Second, we consider a more ideal scenario expecting quantitative distributions of protein concentration. In each, we rank experiments by their utility in discriminating among models and contrast with a ranking focused on refining parameter estimates. In all cases, we correlate utility with variance of the predicted observations, either among models or Representatives, as greater differences are more likely to be discernible. This is an approach implemented previously [36] , [37] , also known as a Maximally Informative Next Experiment [38] and satisfying G-optimality [39] . To illustrate the rankings, the top experiments in each design are presented by heatmaps in Figure 7 , color intensity indicating the relative information gain expected, based on the objective (e.g. variance with parameters, for reducing uncertainty). Refer to Methods for details on the experiment design procedure and calculation of objectives. For each design, the landscape of objective values over all of the experiments considered exhibits a sharp peak, indicating the importance of carefully selecting the experiment (Supporting Information, Figures S15–S17 in Text S1 ).
A) Heatmaps showing relative information gain expected from qualitative data, with experiments on the ordinate and species to measure on the abscissa. Darker boxes indicate greater information (i.e. a more preferable experiment), via expected prediction variance among models (upper), dissimilarity of Representative prediction distributions (center), or variance among Representatives (lower). B) Heatmaps showing experiment design for quantitative data. Based on local sensitivity to a parameter affecting the indicated system feature (ordinate), and ranked by variance among models of mean sensitivity (upper) or variance among Representatives (lower).
https://doi.org/10.1371/journal.pcbi.1003498.g007
Selected experiments from the designs for qualitative data are shown in Figure 8 , where upper panels display expected qualitative predictions and lower plots provide predictions from all Representatives for each model, normalized for visibility. Note that these experiment designs represent a limited range of feasible experiments in this system. More exhaustive model based experiment design carries the promise of more finely resolving system function (e.g. by considering experiments beyond basic genetic perturbations), but is beyond the scope of the current work.
Example simulated results for recommended experiments, showing for each model: qualitative interpretations (upper) and quantitative model outputs for all Representatives, normalized by the mean value to visualize all curves (lower). A) Simulations for experiments recommended directly for model discrimination. B) Simulations for an experiment recommended to first refine acceptable parameter estimates in each model.
https://doi.org/10.1371/journal.pcbi.1003498.g008
Experiment design for qualitative data emphasizes regulators in double mutants.
To compare experiments for qualitative data, we simulated each experiment and translated outputs into an expected qualitative observation of high or low concentration. We included single and pairwise combinations of mutation, heterozygous mutation, and doubled genomic content (indicated by 2×) for each of the proteins common to all networks: Dpp, Receptors (Rec), Mad, Bam, Nos, Brat, and dMyc, a total of 210 experiments.
To evaluate model discrimination power, we first ranked experiments based on their expected qualitative prediction ( Figure 7A , upper heatmap). The expected prediction is the most likely observation predicted by a model, with Representatives weighted equally. For these binary predictions, this corresponds to the median value. Figure 8A left shows the predictions for an experiment measuring pMad in Nos −/− Rec +/− (3rd in upper design of Figure 7A ). While the expected predictions (upper panel) and the mean predictions (lower plots, black lines) vary somewhat among models, much more variance is evident among the Representatives (gray lines) for each model. We expect the experiment to refine parameter estimates, but not clearly discriminate among models. The two best valued experiments return predictions of uniform distributions of Brat and dMyc in 2×Brat Bam −/− and dMyc +/− Nos −/− respectively, varying only in the predicted level. However, the levels predicted for the Brat experiment also vary more widely within each model than among models, indicating that the experiment is unlikely to discriminate clearly. Conversely, the levels for the dMyc experiment are predicted to be consistently high in all models, except Endo where they are variable and low. Accordingly, this experiment is expected to distinguish the Endo model from the rest, but provide no information on the others. Predictions for these two experiments are available in Supporting Information (Figure S18 in Text S1 ).
A drawback to considering expected predictions is that they do not consider how individual predictions are distributed , or how much they overlap among models. To explicitly consider the overlap of prediction distributions , we employ the Jaccard index, which measures the similarity in membership of two sets [1] , [40] . It is defined in a pairwise fashion, so we rank experiments based on the sum of all pairwise indices among models. A zero rank would indicate that all models predict the same set of outcomes, while the maximum value would mean that all model prediction sets are mutually exclusive. This design ranks experiments differently than by expected predictions, though some experiments appear in the top set of both ( Figure 7A , center). The top experiment, measuring Bam in 2×Dpp Nos −/− (appearing 6th for design by expected prediction), is shown in Figure 8A right. Predictions still vary widely within each model. The expected predictions are less diverse than in the design focused on them (upper heatmaps), and the groups of predictions appear more diverse among models (lower plots, gray lines). Examining the prediction distributions however, we expect little clear discrimination except between Alt1 and the rest (i.e. adding support for the Core feedback element).
As the predicted power to discriminate among models is limited by the uncertainty within each model, better identifying parameters for each model should improve the ability to discriminate. To this end, we design to reduce parameter uncertainty by evaluating the variance among predictions from the Representatives in each model ( Figure 7A , lower). The top experiment corresponds with that from the expected predictions (Brat in 2×Brat Bam −/−). Predictions for the 2nd experiment, Nos in Nos +/− Dpp +/−, show wide variance among Representatives, with little difference among models in either mean values or distributions ( Figure 8B ).
Based on these predicted simulations, we expect that a combination of experiments designed to reduce parameter uncertainty and to subsequently discriminate will be most effective. While beyond the scope of this work, design for parallel experiments is a promising approach to more reliably estimate the best set of experiments without performing each sequentially [37] , [41] . For initial experiments, we recommend working to refine model parameters. However, we caution that predictions for spatially uniform data are subject to uncertainty in the sensitivity of the assay used. Accordingly, we recommend experiments predicted to produce non-uniform results, such as measuring Nos in Nos +/− Dpp +/−. It is also worth considering that the measurement of Bam in 2×Dpp Nos −/− and of dMyc in dMyc +/− Nos −/−, as they are expected to test the Alt1 and Endo models, respectively ( Figure 8A right, and S18 in Text S1 ).
Experiment design for quantitative data emphasizes Dpp.
To evaluate experiments anticipating quantitative data, we took a classical approach [14] and evaluated the local sensitivity of model outputs to parameters, for each Representative point. However, our approach differs from classical Fisher Information Matrix based optimal designs in that we consider either model discrimination or uncertainty among multiple Representatives, rather than uncertainty around a single parameter set. We define experiments as the choice of an output to measure and a perturbation related to a model parameter. As with the designs for qualitative data, we evaluate experiments both for discrimination and to refine parameters. To assess discrimination, we rank experiments by the variance of mean sensitivity among models ( Figure 7B , upper). For parameter refinement, we calculated the variance of sensitivity over Representatives, summing over models ( Figure 7B , lower). Recognizing that the model parameters may be affected in multiple ways in the real system, the experiments are listed by the general model feature that is perturbed.
Both designs emphasize measurement of Dpp concentration, which is not expected to be informative in the design for qualitative data. Notably, pMad is the next most useful measurement predicted to refine parameters, and should be considered as well. The two designs differ only slightly in the rank of experiments, indicating little difference between refining parameters and discriminating models.
In this case, because we have employed local sensitivity analyses, we are designing for experiments that perturb the system only slightly. While desirable to limit side effects, this is difficult to implement for internal components in most biological systems. To robustly design for experiments that more significantly perturb conditions, more explicit predictions may be simulated, as with the qualitative design. It is also important to note that we have not considered expected experimental error (i.e. if the predicted results would be distinguishable from the noise). There are several alternative approaches to model based experiment design which may be applicable, depending on the scope and state of the model. For more detail and instruction in experiment design for complex systems, we recommend recent reviews and contributions [41] – [43] .
In considering all designs, it is important to also consider the feasibility of experiments, and any alternative means of acquiring similar data. For these example designs, we have included perturbations of all major system components despite the fact that some may be difficult to produce or to evaluate in a real organism. If such experiments are ranked highly, alternative experiments may be necessary to more practically deliver similar information. For example, some genetic mutants may be lethal or may severely disrupt organism development. However, site specific recombination methods or clonal mutation may be able to provide the relevant information without affecting the entire system as drastically. In such cases, it is also important to properly represent the conditions of the experiment, so models may need to be adapted accordingly.
Conclusions
In this study we have presented a quantitative model analysis based on qualitative data, via multi-objective optimization with Optimal Scaling fitness estimates. Through our analysis of stem cell regulation in the Drosophila germarium, we have demonstrated the estimation of a set of representative parameter sets, discrimination among multiple models, and model-based experiment design.
Using the newly developed process to study the germarium, we have shown the extent to which the existing data employed can discriminate among hypothetical regulatory mechanisms. Current qualitative mRNA and protein image data support the serial inhibition of the (previously presented) Core network, but not the feedback element, which is well evidenced in biochemical data. These data do not distinguish among the more complex mechanisms proposed. Toward future modeling, we indicated data that have yet to be satisfied, model parameters that influence fitness, and presented an example experiment design to improve model discrimination. Based on the limited discrimination expected in the experiment designs performed, we recommend first aiming to reduce parameter uncertainty, e.g. by measuring Nos in Nos +/− Dpp +/−. We also recommend pursuing quantitative measurements for Dpp or pMad, as feasible. The designs presented also indicate a variety of other potential experiments. Beyond these initial experiments however, we recommend a more thorough experiment design with careful attention to the feasibility and cost of different experiments.
The framework we have developed offers benefits in a wide range of applications. In principle, it is appropriate for any mathematical modeling problem where some or all data are limited to qualitative observations. Naturally, there is particular potential for gains in biological applications, where highly complex systems are prevalent. With the Drosophila germarium as a prime example, developmental biology presents many potential applications as it focuses on pattern formation and spatio-temporal behavior, as in the organization of body axes, limbs, and organ structures [44] . In the broader context of biology and medicine, a variety of fields exhibit similar problems and may also benefit from more widespread use of qualitative data in mathematical modeling studies such as this one. General examples include mechanobiology [45] , neurobiology [46] , [47] , and tissue engineering [48] . We would like to emphasize that the techniques developed in this work accommodate uncertainty in data. If all data can be taken in a rigorously quantitative format, the Optimal Scaling procedure is unnecessary. We anticipate that these techniques will be most valuable when including historical data and when employing new measurements that are not yet refined enough to ensure quantitative reporting.
Data compilation
The aggregate dataset of observations on the anterior germarium was assembled from published literature only. Sources were identified by a primary search of combinations of the terms Drosophila, germarium, GSC, bam, brat, nos, and mad. Searches were performed via the search engine Google Scholar and the databases Medline, PubMed, and Science Citation Index. A secondary search identified additional data sources from references within and articles citing the primary findings. Sources were screened for experiments and relevant data.
Data are recorded under a variety of conditions, including genetic mutation and overexpression. Some data were excluded to limit the computational cost of simulations, especially from overexpression studies (e.g. expression via the yeast Gal4-UAS system [49] ) where the increase of expression over wild type is highly uncertain and requires optimization of experimental parameters. Qualitative data were defined by subjective (visual) review of figures and by the interpretations presented by the original authors (e.g. pMad expression is ranked high in a region because its image intensity there appears clearly greater than elsewhere in the same image, with deference to any declared observations made in the published text). Data repeated in multiple works were included one time in the aggregate set, as the observation best representing the consensus from the field. Many data were recorded via fluorescent immunochemistry, which can be ratiometric (i.e. linearly related to the protein concentration) and is often used quantitatively after normalization. However, it is important to consider that the quality of data relies on the entire experiment, not just the final measurement type. The linearity of the data, which is required to reliably normalize, cannot be assured without express guarantees both that the experimental reaction steps were designed to preserve a linear relationship and that the images available accurately present the original intensity values. Many of the experiments aggregated for this study employed enzyme linked visualization assays not originally intended for quantitative comparison or modeling, so controls were not presented to ensure that the reactions remained linear. In addition, the germarium is composed of a soft tissue with a high degree of geometric variability between images, limiting the ability to combine multiple images by geometric registration and evaluate measurement uncertainty. Accordingly, all data were treated as ordinal, which reflects the subjective evaluations presented in the source literature.
To correlate the Phenotype data to Brat expression, we evaluate the mean Brat concentration over the past 6 hours (expecting unmodeled delays, and a cell cycle less than 24 hours [50] , [51] ). Accordingly, data observed with a fusome are assigned a higher rank than those with a spectrosome.
Mathematical modeling
Models of the anterior germarium were designed to represent the system as presented in Figure 1A (see Figure S1 in Supporting Information, Text S1 ). The models consider secretion of Dpp into the extracellular space, diffusion, receptor binding, and protein levels within each cell, according to the internal regulatory network. Alternative models only differ in the intracellular regulatory network, with the exception that the Diff model includes a secreted molecule not modeled otherwise.
Assumptions and implementation.
To form the simplest models appropriate for the system and available data, we apply a set of general assumptions, including: (1) well-mixed conditions, (2) simple saturating regulation, (3) cofactor sufficiency, and (4) one-dimensional organization. 1) Solutions within and near each cell are assumed to be homogenous, i.e. that local diffusion is sufficiently fast for the apparent reaction concentration to be equivalent to bulk concentration. Long-range diffusion, over multiple cell diameters, is explicitly modeled. 2) All protein production regulatory processes, both transcriptional and translational, are approximated by a Hill equation with a cooperativity coefficient of two. The Hill equation provides saturating effects scalable by the half-maximal concentration of the repressor. 3) All cofactors required for reactions are assumed to be present and non-limiting. 4) Geometric effects in directions other than along the anterior-posterior (long) axis of the germarium are assumed to be negligible, given the quality and resolution of the available data.
To implement numerically, the ODEs are coded and solved in MATLAB, using the built-in stiff solver ‘ode15s’. For steady state solutions, ODEs are solved from null (zero value) starting conditions over a simulation period of 24 hours. For dynamically constrained solutions, ODEs are first solved for steady state; then a cell cycle and displacement event is approximated and the results used as initial conditions for a 12 hour simulation. Cell cycle and displacement are approximated by setting each zone (GSC, CB, Cyst, Posterior) to the average solved conditions of its anterior neighbor (i.e. shifting values posteriorly by one zone).
Alternative network identification.
To naively choose networks, we performed two parameter screens for networks that satisfy the available Wild Type data. As all Core regulation is inhibitory, the first screen considered only inhibitory interactions, and was performed by exhaustively sampling combinations of strong and weak regulatory parameters for all possible interactions, excluding self-regulation. Second, to address positive feedback, we performed global optimization over a full range of negative and positive feedback interactions. Both screens were filtered by a fitness threshold and the remaining networks were filtered for parsimony. Further parameter screening details are available in Supporting Information ( Text S1 ).
Optimal scaling fitness estimation
Constraints and implementation.
Numerical procedures
Single objective, global optimization..
To screen for the semi-deterministic global optimization, we allocate samples in a deterministic sparse grid (Chebyshev-Gauss-Lobatto node distribution) [56] , and pseudo-randomly through a latin hypercube design. The sparse grid provides some sampling uniformity and is also used to define a rough polynomial interpolant, which we use to estimate search start points for multiobjective optimization. Bounded by the computational cost of simulating the model, 500 k samples were evaluated for each screen. Sparse grid density was dependent on the size of the parameter space and was chosen to allocate no more than 75% of the samples deterministically. Gradient searches used the MATLAB built-in constrained optimization routine fmincon, via the interior point method, chosen for strict respect of parameter boundaries. 64 gradient searches were run for each single objective optimization.
Multi-objective optimization.
Local sensitivity analysis.
Supporting Information
Dataset s1..
Core model representative parameters. Representative parameter values determined for the Core model, corresponding with Pareto points plotted in Figure 3A , 5A and 6B .
https://doi.org/10.1371/journal.pcbi.1003498.s001
Dataset S2.
Alt1 model representative parameters. Representative parameter values determined for the Alt1 model, corresponding with Pareto points plotted in Figure 5A and 6B .
https://doi.org/10.1371/journal.pcbi.1003498.s002
Dataset S3.
Endo model representative parameters. Representative parameter values determined for the Endo model, corresponding with Pareto points plotted in Figure 6B .
https://doi.org/10.1371/journal.pcbi.1003498.s003
Dataset S4.
Piwi model representative parameters. Representative parameter values determined for the Piwi model, corresponding with Pareto points plotted in Figure 6B .
https://doi.org/10.1371/journal.pcbi.1003498.s004
Dataset S5.
Ago model representative parameters. Representative parameter values determined for the Ago model, corresponding with Pareto points plotted in Figure 6B .
https://doi.org/10.1371/journal.pcbi.1003498.s005
Dataset S6.
Diff model representative parameters. Representative parameter values determined for the Diff model, corresponding with Pareto points plotted in Figure 6B .
https://doi.org/10.1371/journal.pcbi.1003498.s006
Supporting Information. Supporting information regarding model development and analysis.
https://doi.org/10.1371/journal.pcbi.1003498.s007
Acknowledgments
We extend our thanks to Hilary Ashe for conversations and comments on the manuscript.
Author Contributions
Wrote the paper: MP AER DMU. Revised manuscript: MP AER GTB DMU. Conceived and designed procedures: MP AER GTB DMU. Performed modeling and analysis: MP.
- View Article
- Google Scholar
- 11. De Weck OL (2004) Multiobjective optimization: History and promise. In: Invited Keynote Paper, GL2-2, The Third China-Japan-Korea Joint Symposium on Optimization of Structural and Mechanical Systems, Kanazawa, Japan. volume 2.
- 44. Wolpert L, Tickle C, Jessell TM, Lawrence P, Meyerowitz E, et al. (2002) Principles of development, volume 3. Oxford University Press Oxford.
- 56. Klimke A Sparse grid surrogate functions for nonlinear systems with parameter uncertainty. In: Proceedings of the 1st International Conference on Uncertainty in Structural Dynamics. pp. 159–168.

An official website of the United States government
The .gov means it’s official. Federal government websites often end in .gov or .mil. Before sharing sensitive information, make sure you’re on a federal government site.
The site is secure. The https:// ensures that you are connecting to the official website and that any information you provide is encrypted and transmitted securely.
- Publications
- Account settings
- My Bibliography
- Collections
- Citation manager
Save citation to file
Email citation, add to collections.
- Create a new collection
- Add to an existing collection
Add to My Bibliography
Your saved search, create a file for external citation management software, your rss feed, qualitative study, affiliations.
- 1 University of Nebraska Medical Center
- 2 GDB Research and Statistical Consulting
- 3 GDB Research and Statistical Consulting/McLaren Macomb Hospital
- PMID: 29262162
- Bookshelf ID: NBK470395
Qualitative research is a type of research that explores and provides deeper insights into real-world problems. Instead of collecting numerical data points or intervening or introducing treatments just like in quantitative research, qualitative research helps generate hypothenar to further investigate and understand quantitative data. Qualitative research gathers participants' experiences, perceptions, and behavior. It answers the hows and whys instead of how many or how much. It could be structured as a standalone study, purely relying on qualitative data, or part of mixed-methods research that combines qualitative and quantitative data. This review introduces the readers to some basic concepts, definitions, terminology, and applications of qualitative research.
Qualitative research, at its core, asks open-ended questions whose answers are not easily put into numbers, such as "how" and "why." Due to the open-ended nature of the research questions, qualitative research design is often not linear like quantitative design. One of the strengths of qualitative research is its ability to explain processes and patterns of human behavior that can be difficult to quantify. Phenomena such as experiences, attitudes, and behaviors can be complex to capture accurately and quantitatively. In contrast, a qualitative approach allows participants themselves to explain how, why, or what they were thinking, feeling, and experiencing at a particular time or during an event of interest. Quantifying qualitative data certainly is possible, but at its core, qualitative data is looking for themes and patterns that can be difficult to quantify, and it is essential to ensure that the context and narrative of qualitative work are not lost by trying to quantify something that is not meant to be quantified.
However, while qualitative research is sometimes placed in opposition to quantitative research, where they are necessarily opposites and therefore "compete" against each other and the philosophical paradigms associated with each other, qualitative and quantitative work are neither necessarily opposites, nor are they incompatible. While qualitative and quantitative approaches are different, they are not necessarily opposites and certainly not mutually exclusive. For instance, qualitative research can help expand and deepen understanding of data or results obtained from quantitative analysis. For example, say a quantitative analysis has determined a correlation between length of stay and level of patient satisfaction, but why does this correlation exist? This dual-focus scenario shows one way in which qualitative and quantitative research could be integrated.
Copyright © 2024, StatPearls Publishing LLC.
PubMed Disclaimer
Conflict of interest statement
Disclosure: Steven Tenny declares no relevant financial relationships with ineligible companies.
Disclosure: Janelle Brannan declares no relevant financial relationships with ineligible companies.
Disclosure: Grace Brannan declares no relevant financial relationships with ineligible companies.
- Introduction
- Issues of Concern
- Clinical Significance
- Enhancing Healthcare Team Outcomes
- Review Questions
Similar articles
- Folic acid supplementation and malaria susceptibility and severity among people taking antifolate antimalarial drugs in endemic areas. Crider K, Williams J, Qi YP, Gutman J, Yeung L, Mai C, Finkelstain J, Mehta S, Pons-Duran C, Menéndez C, Moraleda C, Rogers L, Daniels K, Green P. Crider K, et al. Cochrane Database Syst Rev. 2022 Feb 1;2(2022):CD014217. doi: 10.1002/14651858.CD014217. Cochrane Database Syst Rev. 2022. PMID: 36321557 Free PMC article.
- Macromolecular crowding: chemistry and physics meet biology (Ascona, Switzerland, 10-14 June 2012). Foffi G, Pastore A, Piazza F, Temussi PA. Foffi G, et al. Phys Biol. 2013 Aug;10(4):040301. doi: 10.1088/1478-3975/10/4/040301. Epub 2013 Aug 2. Phys Biol. 2013. PMID: 23912807
- The future of Cochrane Neonatal. Soll RF, Ovelman C, McGuire W. Soll RF, et al. Early Hum Dev. 2020 Nov;150:105191. doi: 10.1016/j.earlhumdev.2020.105191. Epub 2020 Sep 12. Early Hum Dev. 2020. PMID: 33036834
- Invited review: Qualitative research in dairy science-A narrative review. Ritter C, Koralesky KE, Saraceni J, Roche S, Vaarst M, Kelton D. Ritter C, et al. J Dairy Sci. 2023 Sep;106(9):5880-5895. doi: 10.3168/jds.2022-23125. Epub 2023 Jul 18. J Dairy Sci. 2023. PMID: 37474366 Review.
- Participation in environmental enhancement and conservation activities for health and well-being in adults: a review of quantitative and qualitative evidence. Husk K, Lovell R, Cooper C, Stahl-Timmins W, Garside R. Husk K, et al. Cochrane Database Syst Rev. 2016 May 21;2016(5):CD010351. doi: 10.1002/14651858.CD010351.pub2. Cochrane Database Syst Rev. 2016. PMID: 27207731 Free PMC article. Review.
- Moser A, Korstjens I. Series: Practical guidance to qualitative research. Part 1: Introduction. Eur J Gen Pract. 2017 Dec;23(1):271-273. - PMC - PubMed
- Cleland JA. The qualitative orientation in medical education research. Korean J Med Educ. 2017 Jun;29(2):61-71. - PMC - PubMed
- Foley G, Timonen V. Using Grounded Theory Method to Capture and Analyze Health Care Experiences. Health Serv Res. 2015 Aug;50(4):1195-210. - PMC - PubMed
- Devers KJ. How will we know "good" qualitative research when we see it? Beginning the dialogue in health services research. Health Serv Res. 1999 Dec;34(5 Pt 2):1153-88. - PMC - PubMed
- Huston P, Rowan M. Qualitative studies. Their role in medical research. Can Fam Physician. 1998 Nov;44:2453-8. - PMC - PubMed
Publication types
- Search in PubMed
- Search in MeSH
- Add to Search
Related information
- Cited in Books
LinkOut - more resources
Full text sources.
- NCBI Bookshelf

- Citation Manager
NCBI Literature Resources
MeSH PMC Bookshelf Disclaimer
The PubMed wordmark and PubMed logo are registered trademarks of the U.S. Department of Health and Human Services (HHS). Unauthorized use of these marks is strictly prohibited.
An official website of the United States government
The .gov means it’s official. Federal government websites often end in .gov or .mil. Before sharing sensitive information, make sure you’re on a federal government site.
The site is secure. The https:// ensures that you are connecting to the official website and that any information you provide is encrypted and transmitted securely.
- Publications
- Account settings
Preview improvements coming to the PMC website in October 2024. Learn More or Try it out now .
- Advanced Search
- Journal List
- CBE Life Sci Educ
- v.20(4); Winter 2021
Participation in Biology Education Research Influences Students’ Epistemic Development
† BSCS Science Learning, Colorado Springs, CO 80918
Mallory Wright
‡ Department of Biological Sciences, Clemson University, Clemson, SC 29631
Courtney Faber
§ Cook Grand Challenge Honors Program, Tickle College of Engineering, University of Tennessee, Knoxville, TN 37996
Cazembe Kennedy
∥ Office of Teaching Effectiveness and Innovation, Clemson University, Clemson, SC 29634
Dylan Dittrich-Reed
Knowledge construction is an essential scientific practice, and undergraduate research experiences (UREs) provide opportunities for students to engage with this scientific practice in an authentic context. While participating in UREs, students develop conceptualizations about how science gathers, evaluates, and constructs knowledge (science epistemology) that align with scientific practice. However, there have been few studies focusing on how students’ science epistemologies develop during these experiences. Through the analysis of written reflections and three research papers and by leveraging methods informed by collaborative autoethnography, we construct a case study of one student, describing the development of her science epistemology and scientific agency during her time participating in a biology education URE. Through her reflections and self-analysis, the student describes her context-dependent science epistemology, and how she discovered a new role as a critic of scientific papers. These results have implications for the use of written reflections to facilitate epistemic development during UREs and the role of classroom culture in the development of scientific agency.
INTRODUCTION
As students enter professional careers, they will need to apply their understanding of science to new contexts and construct new knowledge to solve complex problems. To prepare students for such careers, policy makers have highlighted the need to steer student learning toward an understanding of how scientific knowledge is constructed and what counts as knowledge in science, also known as science epistemology ( National Research Council, 2007 , 2013 ; American Association for the Advancement of Science, 2011 ). To pursue the goal of developing students’ science epistemologies, we must first understand epistemic development in students as they participate in authentic science experiences ( Sandoval, 2012 ).
One example of these authentic science experiences are undergraduate research experiences (UREs), in which students engage with research practices to use data and evidence to construct new knowledge within a specific scientific field ( National Academies of Sciences, Engineering, and Medicine [NASEM], 2017 ). There is extensive work describing science, technology, engineering, and mathematics (STEM) student gains in understanding the process of science while participating in UREs (e.g., Thiry et al. , 2005 , 2012 ; Lopatto, 2004 , 2007 ; Hunter et al. , 2007 ), but there is little work describing what epistemic gains may result from student participation in a URE. There are a variety of UREs, and the quality of the educational experience for the student varies based on the costs, research topic, mentoring, and student expectations of the URE ( NASEM, 2017 ). As such, the types of UREs students participate in likely have an impact on their epistemic gains. UREs that focus on biology education (BioEd UREs) provide a unique opportunity for researchers to study epistemic development in undergraduate researchers, because these experiences allow undergraduate researchers to study how others engage with biology knowledge through the use of authentic research practices. We hypothesize that as undergraduate researchers analyze how other students construct knowledge about biology, there will be opportunities for these undergraduate researchers to reflect upon their own knowledge construction. Through these reflections, undergraduate researchers in BioEd UREs will gain a deeper understanding of biology epistemology.
The goal of this paper is to describe one student’s (M.W.) epistemic development through her participation in a BioEd URE and how these changes manifested in her written course work. Because this paper describes a study within a study, we will specifically refer to the URE in which M.W. was an undergraduate researcher as the “BioEd URE,” and the case study in which we investigate M.W.’s epistemic development as the “case study.” M.W. is a coauthor along with her URE mentors, D.L. and D.D.-R. All authors consented to using their initials throughout the article rather than pseudonyms. We begin with an overview of recent research investigating science epistemology, highlighting key outcomes as well as the research approaches, because this work 1) informed the development of the URE project to which M.W. contributed and 2) provides a framework to explore M.W.’s epistemic development as she participated in the URE. Next, we provide a description of the URE project to give context to M.W.’s experience. Then, we present a discussion of M.W.’s experience and evolving science epistemology, taking an approach informed by collaborative autoethnography in which M.W. provides a response to the analysis conducted by D.L. and D.D.-R. Finally, we conclude with a discussion of the implications of this work for future research on science epistemology and approaches to support students’ developing science epistemologies within formal learning environments, such as the classroom and research lab.
BACKGROUND LITERATURE
Supporting the development of students’ science epistemologies.
Epistemology, or the beliefs and approaches around the acquisition, justification, and generation of knowledge, is a core aim of science inquiry ( Longino, 2002 ). Science epistemology establishes the standards for evaluating, justifying, and generating knowledge within science. Students may gain a tacit understanding of epistemology while engaging with the processes of scientific knowledge generation; however, this understanding may be incomplete or inaccurate ( Linn et al. , 2015 ). To ensure that students understand how science generates knowledge, it is important to discuss epistemology while students engage with the process of evaluating, generating, and constructing scientific knowledge ( Sandoval, 2005 ).
Students are exposed to authentic scientific processes during UREs. Many studies have reported that participation in UREs increases student understanding of the processes of science through exposure to authentic scientific practice ( Seymour et al. , 2004 ; Thiry et al. , 2005 , 2012 ; Lopatto, 2007 ; Linn et al. , 2015 ). However, it is unclear whether these experiences help students understand the epistemic foundations of science ( Hunter et al. , 2007 ). Studies investigating the impact of UREs on the development of student epistemology present mixed results. In their review of 53 studies on UREs, Sadler et al. (2010) found that, while some studies reported that students developed an understanding of uncertainty in science and the importance of scientific discourse, other studies reported little or no change in students’ beliefs about how science constructs knowledge.
Practitioners across scientific disciplines from primary school through higher education have implemented classroom interventions to support the development of students’ science epistemologies. The effectiveness of these interventions has been measured quantitatively with Likert-style surveys and qualitatively with open-ended survey items and interviews. For example, in one intervention, undergraduate biology students engaged in analysis of published literature wherein they considered, read, elucidated hypotheses, analyzed and interpreted results, and thought of the next experiment in a process termed C.R.E.A.T.E. ( Hoskins et al. , 2011 ). In a pre–post survey assessment, students rated their own understanding about the nature of scientific knowledge significantly higher in the posttest compared with the pretest ( Hoskins et al. , 2011 ). In another intervention, pre-service elementary school teachers in a geology class participated in a science as storytelling program as a way to teach introductory science students about scientific knowledge ( Bickmore et al. , 2009 ). In this program, students treated science as a form of storytelling with rules that align with scientific practice. Students’ conceptions of science and attitudes toward science were evaluated through surveys that were supplemented by open-ended responses. These pre-service teachers exhibited a better understanding of the creative and tentative aspects of science epistemology and had better attitudes toward science at the conclusion of the course compared with the beginning ( Bickmore et al. , 2009 ). These studies demonstrate the effectiveness of interventions for improving student understanding of science epistemology, but the assessments only report the outcomes of the interventions, leaving us to ask the questions of “how” and “why” students’ epistemic understanding changed.
Several qualitative studies also point to the importance of explicitly discussing epistemology in the classroom for epistemic development. In their study of 8- to 10-year-old children, Ryu and Sandoval (2012) found that students’ epistemologies developed through collective argument, whereby students negotiated epistemic standards for acceptable justifications and appropriated these standards into their argument construction. These results parallel the critical contextual empiricism framework, which describes scientific knowledge construction as a social process whereby standards for knowledge validity are negotiated in a public forum ( Longino, 2002 ). Work by McDonald (2010) points to the importance of explicit instruction in nature of science (NoS) for supporting the development of student understanding of epistemology. During the intervention, pre-service teachers discussed and reflected upon epistemic probes, reflective prompts that directed their attention toward relevant NoS aspects of the lesson ( McDonald, 2010 ). These results suggest that metacognitive tasks such as reflection play an important role in supporting the development of student epistemologies.
Studying Biology Epistemology
The emerging epistemology research in biology education has focused on assessment of the effectiveness of teaching interventions using surveys. Student responses on surveys following the implementation of an active-learning intervention in a large classroom showed that students saw knowledge in biology as a collection of facts transferred from professor to student ( Walker et al. , 2008 ). Supporting this finding are survey results that indicated student perceptions of science epistemology became more novice-like (e.g., memorizing is a primary way of knowing) during an introductory biology class ( Semsar et al. , 2011 ). However, not all assessment of science epistemology resulted in a shift toward novice-like views. Survey results from community college students, first-year students, and advanced students in 4-year colleges exhibited enhanced understanding of science epistemology after exposure to pedagogy involving analysis of scientific literature ( Hoskins et al. , 2011 ; Gottesman and Hoskins, 2013 ; Kenyon et al. , 2016 ). While these survey results present a generalized view of biology students’ epistemologies, qualitative studies present a nuanced view of epistemology that brings context into play.
Surveys inherently assume that student epistemologies exist as coherent cognitive structures that can be accessed through questioning ( Hofer, 2004 ). However, researchers have found that student epistemologies exist instead as a disparate set of resources ( Elby and Hammer, 2001 ; Hammer et al. , 2005 ) that is often tacit ( Hofer, 2004 ). Therefore, surveys, which provide limited opportunity for elaboration, may not capture the nuance and context surrounding students’ perceptions of science epistemology ( Watkins and Elby, 2013 ). Indeed, a qualitative study by Watkins and Elby (2013) focusing on one student’s interview about her views on mathematics in biology found that she held diverse, contextual views about the role of equations in understanding biology.
Qualitative studies in K–12 have made important contributions to our understanding of biology epistemology. For example, researchers who interviewed students between nine and 15 years of age about genetics found that these children’s understanding of genetics consisted of discrete, disconnected units rather than coherent frameworks organized around biological theory ( Venville et al. , 2005 ). This analysis was made possible by the authors’ attention to both the ontological (individual concepts) and epistemological (interconnectedness of the concepts) aspects of genetics understanding ( Venville et al. , 2005 ). The ways in which students unify discrete biological concepts into a coherent framework is also influenced by their learning goals. By studying discourse within a high school classroom, researchers found that students applied different biology concepts to their arguments, in some cases applying these concepts to specifically complete the task at hand (doing the lesson), while in others to gain a deeper understanding of the topic (doing science) ( Jimenez-Aleixandre et al. , 1999 ). These differences in reasoning highlight the importance of students’ goals within particular contexts and their effects on how students apply biological concepts to their epistemic thinking.
Theoretical Framework
Epistemology has been conceptualized by researchers in many different ways: as a set of developmental stages ( Perry, 1990 ; Kuhn, 1991 ; Baxter Magolda, 1992 ; King and Kitchener, 1994 ), a coherent set of beliefs ( Hofer and Pintrich, 1997 ; Schommer‐Aikins et al. , 2005 ) such as the NoS ( Lederman, 2007 ), and as a set of cognitive practices activated in specific contexts ( Louca et al. , 2004 ; Chinn et al. , 2014 ). We chose to conceptualize epistemology as a set of contextual cognitive and metacognitive practices using the epistemic thinking framework ( Barzilai and Zohar, 2014 ), given the findings that student epistemologies are context dependent.
The epistemic thinking framework separates epistemology into two aspects: epistemic cognition (thinking about information) and epistemic metacognition (thinking about knowing). The cognitive aspect is informed by the AIR model for epistemic cognition ( Chinn et al. , 2014 ), which separates epistemology into epistemic a ims, i deals, and r eliable processes to ensure these ideals have been met ( Chinn et al. , 2014 ). “Aims” refer to the objective of the cognitive task, such as determining whether information is accurate ( Chinn et al. , 2014 ). “Ideals” refer to criteria that must be met for an explanation to be accepted as knowledge, for example, ensuring the methods used were appropriate for answering the research question ( Chinn et al. , 2014 ). “Reliable processes” are cognitive practices that are used to achieve epistemic ends (i.e., knowledge or understanding), such as considering multiple perspectives before making a decision ( Chinn et al. , 2014 ). Reliable processes have also been referred to as “epistemic practices” ( Kelly, 2008 ). Taken together, the aims, ideals, and reliable processes of epistemic cognition are the ways that students gather, justify, evaluate, and construct knowledge in a particular discipline like biology.
Epistemic metacognition is an individual’s awareness of the knowledge, skills, and experiences related to that individual’s thinking and learning. Much like metacognition, epistemic metacognition is divided into three subcategories: epistemic metacognitive knowledge (EMK), individuals’ knowledge about how they and others conceptualize knowledge; epistemic metacognitive skills (EMS), the different ways people evaluate, monitor, or plan how to reach an epistemic aims/ends; and epistemic metacognitive experiences (EME), what people are aware of or feel as they are working toward an epistemic aim ( Barzilai and Zohar, 2014 ). Just as metacognition has been shown to affect the way biology students approach learning ( Stanton et al. , 2019 ), we hypothesize that epistemic metacognition will affect the way that students approach scientific knowledge. Using this theoretical framework, we aim to address the following research questions: 1) In what ways does one student’s (M.W.) participation in a biology education research URE affect her epistemic development? 2) How, if at all, are these changes manifested in her written course work?
The goal of our study was to explore M.W.’s epistemic development within the context of a BioEd URE and her biology course work. We used a case study approach combined with M.W.’s autoethnographic descriptions, which allowed us to consider M.W.’s epistemology within the context of the BioEd URE and her biology course work. Through this combination of methods, we present a description of M.W.’s epistemic development, incorporating our analysis of her course work and experiences in the BioEd URE with her own perspective of the experiences.
Study Context: The BioEd URE
M.W. joined a BioEd URE investigating undergraduate biology students’ thoughts about scientific knowledge in the Spring of 2018. In this experience, M.W. was an undergraduate researcher, D.L. was a graduate researcher, and D.D.-R. was the principal investigator. The aim of the BioED URE was to answer the research question: How do students participating in a scientific argumentation–focused introductory biology course construct arguments in a literature review compared with students participating in a lecture-based introductory biology course? As part of the BioEd URE, we collected student research papers from two sections of an introductory biology course and analyzed the papers to identify students’ arguments and reasoning to explore students’ science epistemology. M.W. took this introductory biology course and completed these research paper assignments in the Fall of 2016 and Spring of 2017.
In order for M.W. to effectively analyze the research papers for science epistemology, she needed to be well versed in epistemic theory. As such, D.D.-R. and D.L. included readings, weekly discussion, and written reflections on epistemic theory in M.W.’s BioEd URE. In particular, we assigned M.W. readings on the AIR model for epistemic cognition ( Chinn et al. , 2014 ) and the epistemic thinking framework ( Barzilai and Zohar, 2014 ). Once she was familiar with these theoretical frameworks, M.W. began analyzing participants’ scientific arguments within their course research papers. This analysis included the construction of a codebook through both emergent and a priori coding. Throughout this process, our research team held weekly meetings to discuss general research practices and engage M.W. in reflection on how the epistemic theories related to her own thoughts about scientific knowledge in the context of her experiences. The integration of reflection was informed by the work of Kalman (2007) and was included to support M.W.’s thinking about the epistemic theories we discussed. Over the course of one semester, we asked M.W. to write nine reflections. The specific prompts grew out of the discussions about science epistemology during our lab meetings. In her second reflection, M.W. writes:
When I started this project, the whole idea of epistemic cognition seemed very far-fetched and abstract. I didn’t really understand how it was possible to study such internal thoughts of other people by simply reading their papers. This is still a challenge for me now because I find it hard to put myself in others’ shoes and try to understand their intentions when writing these papers. How can we really find out the truth about how “people know what they know?” This question still stumps me.
When [D.D.-R.] asked me about how I was reacting to trying to understand our research, I told him it was making me second guess my past writing. For example, do I really blindly trust all scientific sources on the internet simply because they are published? And even if and when I do this, does it actually affect my writing on a deeper level?
I decided to skim through my own biology lab [research] papers from last year to see how my own writing compares to the papers that we have been reading and coding thus far. One thing that I noticed about my papers was that I explained a lot of the background information in my own words and used a citation at the end of the paragraph that supported my explanation of the scientific mechanisms. For example, I wrote down the process of the cell cycle and explained it in my own words, then searched for a source that re-iterated what I said in my paper.
These insightful reflections on her own epistemology led us to reorient our research lens onto M.W.’s epistemic development. Consequently, her written reflections became an important part of the data set for the present study.
In addition to carefully designing training around epistemic theories for M.W., we (D.D.-R. and D.L.) also strove to create a community where M.W. felt comfortable challenging our interpretations, which was important to maintaining research quality for the original BioEd URE study. To create this community, we mirrored the four norms of an ideal scientific community outlined by Longino (2002) in her description of critical contextual empiricism:
- Providing venues for criticism gives researchers a place to critique ideas so that only the most well-supported ideas are accepted as knowledge.
- Uptaking criticism allows researchers to evaluate ideas based on criticism and make changes to these ideas when appropriate.
- Recognizing public standards and using these standards to evaluate ideas helps a community maintain the quality of its knowledge.
- Maintaining tempered intellectual equality ensures that voices within the community are heard, while ensuring that the influence of the voices are tempered by each individual’s expertise.
We provided a venue for criticism of ideas in the form of research meetings, where we modeled the uptake of criticism and how to make appropriate changes to data interpretations in response to that criticism. During these research meetings, we also discussed the public standards of research quality in the context of both quantitative biology research and qualitative biology education research. We maintained tempered intellectual equality by considering all ideas presented and explaining our reasoning and theoretical justification when necessary.
All of the aspects described, including the specific training on epistemic theories and the community mirroring Longino’s (2002) four norms, are part of the context under which we (all authors) seek to understand M.W.’s epistemic development. The other part of the context that undergirds M.W.’s BioEd URE experiences is her progression through her biology course work, which is briefly mentioned by M.W. herself in the quote presented earlier. We will provide more details about these courses and the research papers she writes in later sections.
Research Quality Framework
The quality framework (Q3) developed in engineering education ( Walther et al. , 2013 ; Sochacka et al. , 2018 ) provided the language to describe and guide our thinking on key research quality issues throughout our data collection and analysis for our case study. Q3 separates interpretive research quality issues into six constructs: theoretical, procedural, communicative, pragmatic, and ethical validity, and process reliability ( Table 1 ).
The Q3 research quality framework
| Quality construct | Construct definition |
|---|---|
| Theoretical validation | The theory generated from the analysis is representative of the social reality under study. |
| Procedural validation | The research design ensures that knowledge built from the project aligns with the social reality being studied. |
| Communicative validation | The ways in which data and analyses are effectively communicated between members of the research group, discipline, and beyond. |
| Pragmatic validation | The theoretical framework(s) used in this study are compatible with the social reality under investigation. |
| Ethical validation | The ways in which the researchers consider the underlying human elements that govern the influences between researchers and participants. |
| Process reliability | The processes used in this project are dependable and consistent. |
Ethical validation was an especially important aspect of this study because of the inclusion of M.W. as a researcher/participant. The guiding questions presented by Sochacka et al. (2018) shaped our thinking on how to equitably engage M.W. as a researcher, ensure that our analysis did justice to her lived experience, and temper our own biases so that they did not unduly influence M.W. or the interpretations we present. We will use the language described in Table 1 to discuss other affordances and challenges to the aspects of research quality throughout this paper.
Participant as Researcher
Given the nature of this study and to ensure that M.W.’s voice was appropriately represented, the BioEd URE research team (D.D.-R., D.L., and M.W.) contacted their local Institutional Review Board (IRB) for guidance. Following an IRB-approved procedure, M.W. provided written consent to be identified as a researcher participant (IRB approval no. 2016-244). As an identified researcher participant, M.W. contributes her insights throughout this paper, displayed in italics. To address the quality aspects of communicative and theoretical validation, and to ensure that her voice is preserved, we elected to keep her commentary separate rather than incorporating her comments into the narrative of the paper. As such, “we” represents the combined voices of D.L., D.D.-R., C.K., and C.F. This way, readers can differentiate between the researchers’ analyses and can experience M.W.’s self-analysis in her own words. As a part of ethical validity, each author is referred to by initials in this paper to maintain intellectual equality among the researchers. Each researcher’s involvement in the project is described in Table 2 .
Description of researcher roles on project
| Description | Researchers | |
|---|---|---|
| Coders | Analyzed papers and reflections. Wrote and revised the paper and reflection analysis memos. Constructed themes. | D.L. and D.D.-R. |
| Critical peer review | Read the paper and/or reflection analysis memos. Critiqued the analysis and conclusions. | C.F. and C.K. |
| Autoethnographic review | Critiqued analysis and conclusions and provided thick autoethnographic descriptions of classroom and BioEd URE experiences | M.W. |
Participant Description
At the time of the study, I was a sophomore microbiology major and sociology minor. I was also an honors college student, taking honors biology and chemistry courses at Clemson University. Due to my microbiology major, I took very specific courses on microbes, but also took broader biology courses such as cell biology and immunology. My sociology minor allowed me to take classes about social topics like deviance, drug abuse, and the family. I was not interested in sociology until I came to Clemson and took an introductory sociology class to fulfill a requirement, which inspired me to take more classes. This interest in sociology broadened my interests to include social science in addition to my traditional “hard” science classes (i.e. biology and chemistry).
I previously participated in undergraduate research my freshman year. I worked in a life sciences lab and learned basic skills, such as how to grow cells in culture and count cells accurately, in order to design and implement my own experiment. The experiment I worked on consisted of investigating the effects of fruit and vegetable extracts on cancerous cells. Additionally, I was a biology peer mentor for the first semester of my sophomore year, which introduced me to the Engineering and Science Education department. I then joined this project [BioEd URE] and participated in another form of undergraduate research. In some ways, I am a typical microbiology major: I am on the pre-med track and interested in the public health side of microbiology. However, my interest in sociology makes me different from others in my major because these subjects don’t always cross paths past the introductory sociology requirement. Also, I worked with students as an Orientation Ambassador and a biology peer mentor, so I am interested in learning more about the education aspect of biology and learning more about how students like myself learn about biology.
Participant Curricula during the BioEd URE
In addition to the general participant description M.W. provided, we further contextualize her experience by describing some of the course work she completed concurrently with the BioEd URE. M.W. participated in the BioED URE for one semester and was not able to continue the project because of curricular and time constraints. During the BioEd URE semester, M.W. was enrolled in 11 credits of science courses, a 3-credit psychology course, a 3-credit science writing course, and the 2-credit BioEd URE, for a total of 19 credits.
The science writing course likely affected M.W.’s writing skills, so we present some details about this course, beginning with the course description.
[This Science Writing Course] introduces students to the study and practice of professional scientific communication through the analysis of and writing of the major genres in the discipline. It focuses on the principles, strategies, and styles of scientific argumentation and audience adaptation in written media. It is designed for students in the sciences.
As part of the course, M.W. completed a literature review paper. We present the rubric for the literature review assignment in Appendix C in the Supplemental Material. In particular, criteria 4 and 7, emphasizing synthesis of research articles and constructing your own conclusions could have been influential in M.W.’s writing for the literature review assignment.
Research Design
The contextual nature of epistemic cognition ( Hammer et al. , 2005 ; Watkins and Elby, 2013 ; Chinn et al. , 2014 ) compelled us to study M.W.’s science epistemology in context. Given our focus on context, we chose to construct a case study that “investigates a contemporary phenomenon (the ‘case’) in depth and within its real world context, especially when the boundaries between phenomenon and context may not be clearly evident” ( Yin, 2018 , p. 15). Aligning our study with a case study approach also provided a means to ensure procedural validity through the general methodology provided by this approach. A case study approach is a flexible methodology that can accommodate a variety of data sources ( Baxter and Jack, 2008 ), which allows us to leverage research papers and written reflections generated by M.W. to produce a thick description of her case. These descriptions allow researchers to answer “how” and “why” questions about phenomena over which they have little or no control ( Yin, 2018 ), such as how or why student epistemologies developed in response to an intervention. In fact, case study methodology has been used by researchers to study science identity ( Tan and Barton, 2008a , b ) and science epistemology ( Watkins and Elby, 2013 ).
Despite the benefits of case studies, some researchers express concerns around the scope, rigor, and generalizability of the results. Case studies generate a vast pool of data, which may tempt researchers to answer questions that are too broad. To address this constraint, it is important that case studies are bound by time, place, or context ( Stake, 2006 ; Creswell, 2012 ; Yin, 2018 ) and that the researchers define the unit of analysis to focus on salient parts of the data ( Baxter and Jack, 2008 ). This case study is bound by time and context. The analysis is bounded by time in the sense that the analysis focused on the time M.W. spent as a researcher in the BioEd URE. To provide more context about the development of her thinking about scientific knowledge, we also analyzed assignments she completed 1 year before the BioEd URE (research papers she had previously written), and one semester after the BioEd URE (a reflection she wrote about the BioEd URE after the experience had concluded). The unit of analysis is M.W. herself. Finally, case studies “are generalizable to theoretical propositions and not to populations or universes” ( Yin, 2018 , p. 20). In other words, our case study results can be used to expand epistemic theory, but not to extrapolate the behavior of students outside our case. To consider the case in light of other students, Stake (2000) suggests that researchers “describe the cases in sufficient descriptive narrative so that readers can vicariously experience these happenings and draw conclusions (which may differ from those of the researchers)” (p. 439). To ensure the transferability of our case study to other contexts, we provide descriptions that faithfully represent M.W.’s lived experience. To ensure the authentic representation of M.W.’s lived experience, we combined our case study approach with elements of an autoethnography.
Autoethnography is a research approach that combines elements from autobiography and ethnography, allowing researchers to explore a cultural phenomenon through their own personal experiences ( Ellis et al. , 2011 ; Hughes et al. , 2012 ). Autobiography describes events that led to significant change in the author’s life, and ethnography explains how engagement with a culture made these moments of change possible ( Ellis et al. , 2011 ). Within autoethnography, it is important that the personal experiences, thoughts, and actions are documented and made visible for analysis. Additionally, it is important that the researcher moves from experience-near (their own experiences) to experience-far (larger cultural relevance) throughout data collection and analysis. There are multiple approaches that can be used to support this process. For this work, we used M.W.’s responses to the URE reflection prompts and our research team discussions. These data were analyzed by experts within the theoretical space. The reflection prompts and research team discussions supported M.W.’s documentation of her own personal experiences, thoughts, and actions, making them visible for analysis and providing her with the space to consider her own context. D.L. and D.D.-R. developed the reflection prompts and participated in the research team discussions, providing a means to support the process of going from experience-near to experience-far. Specifically, they were able to ask additional questions of M.W., allowing further exploration of specific experiences, and they were also able to guide her developing understanding of epistemic theories, allowing M.W. to participate in the process of analyzing her own experience and reflect specifically on her developing epistemic cognition. Much of the initial data analysis was conducted by D.D.-R. and D.L. because of their expertise and understanding of epistemic cognition; however, M.W. was actively engaged in data analysis through extensive member-checking, reviewing, and providing feedback on D.D.-R. and D.L.’s analysis. This process ensured that the outcomes of this work provide an authentic representation of M.W.’s experience and go beyond M.W.’s own experience to make larger statements on the general cultural phenomenon of developing students’ epistemic cognition. It is through the combination of our case study analysis perspective and M.W.’s autoethnographic lens that we seek to explore how M.W.’s engagement in this biology education URE affected her science epistemology.
Qualitative Data Selection
This study grew from discussions with M.W. during her experience with the BioEd URE. Consequently, the data we analyzed were not so much collected, but selected from assignments that M.W. completed during her time as a researcher in the BioEd URE. The data for this study consisted of three papers M.W. wrote for course work and reflections she wrote during the BioEd URE ( Figure 1 ). M.W. wrote the first two papers for an introductory biology class during her first year (academic year 2016–2017): one in the Fall semester and the other in the Spring semester. Both papers were literature reviews on a scientific issue related to biology, referencing peer-reviewed journal articles. The instructions for the assignments were identical, except that students were asked to include an ethics section in the paper in the Spring semester. The rubric for the biology literature review papers can be found in Appendix B in the Supplemental Material. We selected M.W.’s two introductory biology papers because they were part of the data set for the BioEd URE. As we describe in the Results , M.W. had begun a self-analysis of her science epistemology of her own accord, starting with these two papers. It was this self-analysis that inspired the development of the present case study. M.W. wrote the third paper for a science writing course in her sophomore year. This assignment was also a literature review on a scientific subject, and M.W. chose to write the paper on a subject related to biology. We included this paper in the case study, because it provided an opportunity to explore M.W.’s epistemology in a similar context: through her scientific writing in a literature review.

Timeline showing the sequence of M.W.’s courses, the three course research papers, her BioEd URE, the autoethnographic study, and the 10 written reflections. M.W. wrote her first two research papers in an introductory biology class in Fall and Spring semesters of 2016-17. She wrote her third paper in a science writing course in Spring 2018 while she was concurrently participating in the BioEd URE. The autoethnographic study, looking back on her experiences in her science courses and during the BioEd URE, occurred during the Fall.
M.W. wrote a total of 10 reflections, nine written during the URE and the 10th during the Fall semester of her junior year ( Figure 1 and Table 4 ). The nine reflection prompts during the URE were all derived from discussions we had with M.W. during research meetings. The 10th reflection asked M.W. to reflect on her epistemic growth by asking her whether or not she believed she could write a paper of the same quality as her third literature review paper as a first-year student, and if there was anything she would change about the papers she wrote for her introductory biology class. The topic of each of the reflections is stated in Appendix A in the Supplemental Material. No guidance was given on format or length, but M.W. generally kept reflections to one typed page, single-spaced.
BioEd URE reflection questions
| Reflection date | Reflection question | Justification |
|---|---|---|
| January 23 | Reflect on the epistemic aims, ideals, and reliable processes in the context of your classes and in the context of a real-world problem. | This reflection was assigned to help M.W. familiarize herself with the components of the AIR model for epistemic cognition ( , 2014). |
| January 30 | What are you struggling to understand in this research project? How does one know what information to trust or not trust? | In her initial reads through the data, M.W. found it difficult to interpret students’ papers. This reflection was assigned to help her think about what counts as trustworthy information. |
| February 11 | Find an article using Web of Science/ERIC or another database. Summarize and critique the paper. | This reflection was assigned to help M.W. find peer-reviewed articles using a literature database. Summarizing and critiquing the paper was an exercise to help M.W. develop confidence in critiquing published literature. We let M.W. choose her own paper so that she could choose a topic that was most interesting to her. It was important for M.W. to critique literature so that she could find strong articles that were pertinent to the BioEd URE. |
| February 14 | Reflect on how you came up with “fake chemistry” to find a correct answer on your chemistry exam. | M.W. had just taken an exam and felt that she had made up “fake chemistry” to answer a question. This reflection was assigned to help M.W. understand how she selected bits of prior knowledge to construct her answer. We felt that reflecting on this kind of knowledge construction would help her understand how other students might construct knowledge in our data. |
| March 3 | Examine the clarity/correctness framework by Cheatham and Tormala. Can you connect what students are saying to what they know by using this framework? | This reflection was assigned to help us determine whether the clarity/correctness framework was suitable for the BioEd URE data analysis |
| March 7 | In your mind, what is the difference between your experience in a laboratory research experience vs. this education research experience? | This reflection was assigned to help M.W. think about the similarities and differences in epistemologies between different contexts. |
| March 11 | You mentioned that you wrote a literature review for your science writing class. Reflect on how you wrote that literature review and compare it to how you’ve written other literature reviews. | M.W. told us of a literature review she wrote in a science writing course. This reflection was assigned to help M.W. think through how the epistemology she used when writing the literature review was similar to and/or different from the epistemology she used when writing Papers 1 and 2. We reasoned that thinking through differences in how she applied her own epistemology would help M.W. to analyze the BioEd URE data. |
| March 29 | Block off what you believe to be the student arguments in the paper. Once blocked off, reflect on what parts of the student’s paper are important to our analysis. | This reflection was assigned to help M.W. analyze the BioEd URE data. |
| April 4 | You mentioned that one of your friends changed her paper topic because she was afraid that she would not agree with her TA. Think about how you and others choose your paper topics, and whether or not it affects how you look for evidence. | This reflection was assigned to help M.W. think about student motivations and how these motivations might affect their epistemologies. |
| October 7 | (After the research project) Reflect on the two papers you wrote in your introductory biology class and the paper you wrote in your science writing class. Could you have written the paper you wrote in your science writing class as a freshman? What would you change about these papers now? | This reflection was assigned to indirectly ask M.W. how her epistemology may have changed between writing Papers 1, 2, and 3. |
Qualitative Data Analysis
In the present case study, we analyzed M.W.’s three research papers and the 10 reflections she wrote in connection to the BioEd URE ( Figure 2 ). M.W.’s research papers were analyzed for empirical evidence of changes in her science epistemology. Her reflections were analyzed to determine what aspects of her education, which included the BioEd URE, were influential in the development of her science epistemology. Analyses of M.W.’s research papers and reflections were summarized in two analysis memos: one for her research papers and one for her reflections ( Figure 2 ). All data were consensus coded by D.L. and D.D.-R. by first coding the data separately, then meeting to discuss code definitions and meanings. They reconciled disagreements through discussion, applying codes that aligned best with the data.

Summary of analysis. M.W. (Author 2) helped to refine the themes and case descriptions by leveraging her autoethnographic descriptions. C.K. (Author 4) provided a perspective on the analysis that was further removed from the data. D.L. (Author 1) and D.D.-R. (Author 5) were involved throughout the analysis process.
Analysis of the three research papers focused on the claims M.W. presented, the data she used to support the claims, and the warrants that explained the connections between her claims and her data, as described by the Toulmin argument pattern (TAP; Toulmin, 2003 ). We view argument as an epistemic practice, a means by which knowledge is justified ( Kuhn, 1991 ; Kelly, 2008 ). As such, the kinds of data and warrants M.W. employed to support her claims give valuable insight into the ways she thought about knowledge in science. Analysis of M.W.’s research papers began with a read-through to get a feel for the data, followed by coding of the reference section. The coding pair identified arguments using TAP, noting connections between argument structures and/or identifying an overarching argument, if present. These identified arguments were coded, taking into account the kinds of sources M.W. used as data, the ways in which she described the data from the sources, and how she used the data to support her hypothesis. For example, where M.W. restated the conclusions from a particular source, we coded these excerpts as “reporting.” In contrast, where M.W. used data from multiple sources to construct an assertion not found in those sources, we labeled these excerpts as “synthesis.” Once coding was complete, an analysis memo was written to integrate meaning-making from the paper analysis. More details about the analysis memos are provided at the end of this section.
Analysis of M.W.’s reflections focused on her epistemic thinking. Like the research paper, analysis of these data began with a read-through to familiarize ourselves with the data. We then analyzed the data by identifying excerpts related to the epistemic thinking framework ( Barzilai and Zohar, 2014 ). Leveraging the epistemic thinking framework in our coding helped us to identify excerpts that demonstrated M.W.’s EMK about science, and the EMS she used to develop this knowledge. D.L. and D.D.-R. initially planned to code the reflections similarly to the paper analysis, but the first attempts at coding made it evident that deconstruction of the data into constituent parts left many of the details of M.W.’s epistemic development undescribed. To address this challenge to theoretical validity, D.L. and D.D.-R. shifted their approach to one informed by narrative analysis, which allowed them to consider the reflections as a coherent whole ( Polkinghorne, 1995 ). The identified excerpts were grouped in chronological order, and a narrative was written in the form of an analysis memo, using the excerpts from M.W.’s reflections as a framework.
The analysis of the research papers was also summarized in separate analysis memos that were coconstructed by the coders. Both analysis memos included a descriptive representation of the data followed by a summary of salient interpretations emerging from the analysis ( Lee et al. , 2019 ). The analysis memos were written by either D.L. or D.D.-R. Once the analysis memos were drafted, D.L. and D.D.-R. reviewed and revised them until consensus was reached. To enhance theoretical validity, a third researcher, C.K., who did not code the data, critiqued the data and analysis memos written by the coding team by looking for data that contrasted with conclusions drawn by D.L. and D.D.-R. C.K., D.L., and D.D.-R. discussed any disagreements until they reached consensus; then the analysis memos were finalized.
Theme and Narrative Construction
Once the two the analysis memos were finalized, D.L. and D.D.-R. read through them to integrate the paper analysis with the reflection analysis. They then individually generated a list of themes and met to discuss each theme to decide if the themes were salient or should be combined. Once they reached consensus, D.L. and D.D.-R. wrote descriptions of each tentative theme. The themes served as the principal components that facilitated the retelling of how M.W.’s science epistemology developed during her time spent participating in the BioEd URE. At this point, C.K. critiqued the theme descriptions and the narrative, attempting once again to disconfirm each theme. C.K., D.L., and D.D.-R. discussed any disagreements on the theme descriptions and narrative until they reached consensus, then revised the narrative as necessary. Once finished, the theme descriptions and narrative were presented to M.W., who refined the narrative through her autoethnographic lens. M.W. wrote responses to each theme, highlighting points of agreement and disagreement, drawing from her own experience to provide evidence for her claims. The research team (including M.W.) then met to resolve any disagreements and revise the narrative ( Figure 2 ).
Through our analysis of M.W.’s papers and reflections, we tell the story of M.W.’s developing science epistemology, which resulted in her development of agency toward constructing scientific knowledge. The diversity of artifacts that we collected allowed us to assess M.W.’s epistemic practices. The research papers we collected illustrate M.W.’s use of epistemic practices in the context of her classroom experiences ( Table 3 ). It is clear from her research papers that M.W. shifted from listing facts from instructors and peer-reviewed sources to building reasoned arguments of her own making between papers she wrote before and during the research experience.
Research paper analysis summary
| Paper | Paper context | Primary epistemic practice | Representative quote |
|---|---|---|---|
| 1 | Introductory Biology class, Fall of freshman year | Reporting information; no conclusions are made; facts are presented without further explanation. | “Palbociclib is a CDK4/CDK6 inhibitor that as of February 2015, has been approved by the Food and Drug Administration (FDA) toward treating breast cancer. This inhibitor works by targeting and stopping the production of CDK4/6 in cells. This inhibitor dephosphorylates the protein pRb along with arresting the G1 phase of the cell cycle.” |
| 2 | Introductory Biology class, Spring of freshman year | Simple arguments, M.W. attributes her conclusions to her sources. | “Genetically diverse crops differ in that the population is able to resist extreme changes in environmental conditions because some are more resistant than others to changes in environmental factors. The decrease in biodiversity of livestock feed crops is dangerous because it increases the likelihood that the crops will undergo massive crop failure, leading to unprecedented changes in the global food supply (Di Falco, 2004).” |
| 3 | Science Writing class, Spring of sophomore year | Complex arguments, M.W. synthesizes information from multiple sources to construct her own conclusions. | “The current method of treatment includes three therapies that target the bacteria themselves: proton pump inhibitors, amoxicillin, and clarithromycin (Molina-Infante and Gisbert, 2014). However, the efficiency of these antibiotics is on the decline, with studies showing a decrease from 81.3% to 77.5% antibiotic effectiveness (Chung , 2011). Some of the main factors found to contribute to this decrease are antibiotic resistance, drug compliance, and degradation of the antibiotics by the acidic nature of the stomach (Chung , 2011). Therefore, the future for treating H. pylori infections lies in finding more effective antibiotics as well as alternative treatments besides antibiotics.” |
From the analysis of her research papers, it is not clear why M.W. shifted her approach from reporting information to knowledge construction. However, M.W. reveals the reasons for the changes in her biology epistemology through her written reflections. Furthermore, her self-analysis of the data we collected filled many of the gaps left from our analysis. For this reason, we focus our efforts in this paper on the reflections M.W. wrote during the BioEd URE. In the following sections, we tell the story of M.W.’s development of science epistemology through the reflections she wrote during the BioEd URE. We support this narrative with selections from M.W.’s responses to our analysis, presented in italicized text . Through the chronological analysis of M.W.’s reflections, we found that her epistemic development occurred through three distinct steps. First, M.W. realized that her thoughts about knowledge differed between contexts. The realization that her epistemology was situated and differed between contexts allowed her to reflect on her perceptions about her role as someone who could challenge published claims in the context of the BioEd URE. M.W.’s reflections about her ability to challenge published claims influenced her development of agency toward scientific knowledge production. We describe each component of the narrative in greater detail in the following sections.
M.W.’s Thoughts about Knowledge Differ between Contexts
Previous work has found that individuals’ thoughts about knowledge is contextual ( Louca et al. , 2004 ; Chinn et al. , 2014 ), so we begin our description of M.W.’s epistemic practices with a discussion about the contexts in which she places her epistemology. Through her first two reflections, M.W. describes three contexts in which she interacted with knowledge from her own perspective: during an undergraduate science class, while thinking about sociocultural issues, and while citing scientific papers. Upon reflecting on these contexts, M.W. explains how she views and interacts with knowledge within these contexts. At the beginning of the BioEd URE, M.W. makes a clear delineation between her thinking in science class and with sociocultural issues such as making decisions about universal healthcare. Her first written reflection reveals diverging ideas about how she determines what is correct in classroom and sociocultural contexts.
My aim or goal in [STEM] class is to get a good grade so that I can get into a top graduate school program. I determine what is right in class by what my professor says. If he teaches a topic a certain way, I assume that he is right because he is the one that will end up grading my papers.
[…] My aim when evaluating our healthcare system is to learn the truth so that I can make an educated decision on whether I support or do not support universal health care. I want to make an educated decision, rather than just going along with what my friends or family believes.—Reflection 1
These excerpts reveal M.W.’s classroom aim of “getting a good grade” in a STEM class context, and her sociocultural aim to “learn the truth” in the context of making decisions about healthcare policy. She describes a difference in decision making between the two contexts: she defers to the instructor in STEM class but makes her own educated decision when talking about healthcare policy.
In her second reflection, M.W. analyzes her own research papers ( Table 3 ) and reflects on her thinking. We did not ask M.W. to analyze her own research papers as part of the reflection; she decided to do this on her own. The following excerpt is a part of this self-analysis.
I fell into the routine of almost paraphrasing what the articles said, rather than interpreting them myself. I think that I do this because I trust the publications, and since I didn’t do the trials or research on my own, I don’t feel like I am in a position to challenge their claims.—Reflection 2
Through her self-analysis, M.W. finds that she does not feel like she is “in a position to challenge” claims made by researchers, because she “didn’t do the trials or research on my own.” Her perception that she cannot challenge the claims made in publications occurs within a third context, where M.W. feels she is only able to question claims if she was involved in data collection or analysis.
M.W.’s Perception of Her Own Place in Challenging Research Claims Changed during the BioEd URE
During the BioEd URE, we provided M.W. with activities explicitly designed to increase her willingness to challenge scientific claims. We contend that these activities influenced M.W.’s willingness to challenge claims made by scientists. For example, 1 week after we assigned Reflection 2, we discussed the issue of underdetermination, the idea that multiple interpretations can be drawn from the same body of evidence. We used this discussion to stress to M.W. the importance of considering multiple interpretations and forming her own conclusions, even if they differed from ours. Following this discussion, we asked M.W. to find a published journal article and summarize it in a written reflection so that she could practice interpreting data and forming her own conclusions. M.W. read the article she chose with a critical eye.
My issue with this article was that the abstract presented the findings in a confusing way so that after I finished reading the article, I felt like the authors had lied to me. The abstract states, “results indicate that the presentation of controversial topics, particularly evolution, in the context of public health could be used to encourage public acceptance of scientific viewpoints.” However, the discussion/conclusion talks about how the study showed no support of the student’s acceptance of global warming being influenced by evidence-based explanations. The study did show a significant change in the student’s opinions on evolution, but not on global warming. Therefore, the wording of the abstract is misleading because it implies that their theory can be applied to many topics or on a larger scale; this is not necessarily true.—Reflection 3
This excerpt demonstrates M.W.’s ability to critique the claims of researchers and her willingness to do so in the context of the BioEd URE. It was interesting to find M.W. critiquing the claims made by authors of her selected article because of the statements she made in Reflection 2: “I trust the publications, and since I didn’t do the trials or research on my own, I don’t feel like I am in a position to challenge their claims.” The short time between Reflection 2 and Reflection 3 (12 days) suggests that M.W. already possessed the skills to critique scientific literature but did not feel that it was proper for her to form her own conclusions in specific contexts. In the following extract, M.W. explains why she was able to challenge the conclusions made in the published article. The excerpt is M.W.’s self-analysis of her own work, so it is presented in italics.
In the context of the reflection, I was able to challenge the paper because it was my own reflection, there was not a right or wrong answer, and it was solely my opinion. Just like determining my stance on healthcare, it was a place for me to determine my own opinion. In STEM class, there is no room to decide what I think is right or wrong, the subject requires me to learn the processes and present it on the test.
M.W. explains in this self-analysis that the difference in context between the reflection and STEM class facilitated her willingness to challenge claims made in a published journal article. However, there is also evidence that her willingness to challenge scientific claims made in published literature transferred to the paper she wrote in her science writing course (Paper 3). In the following excerpt, M.W. critiques the claims made in a paper describing antibiotic treatment regimen.
One newly developed antibiotic treatment developed in 2000 is called sequential therapy. This therapy treatment includes a proton pump inhibitor (PPI) and amoxicillin for 5 days, as well as a PPI, clarithromycin, and tinidazole triple therapy for an additional 5 days. This treatment method was found to have a higher eradication rate than the standard triple therapy described previously. This higher rate was contributed to the decreasing H. pylori density in the stomach and corresponding increase in the effectiveness of the antibiotics clarithromycin and metronidazole. 16 However, these studies fail to investigate whether the improvement in the eradication rate is due to the sequential therapy or the increased amount of antibiotic use.—Paper 3
As in her first and second research papers, M.W. cites scientific journal articles to support her claims. However, unlike in her first two papers, M.W. qualifies data presented by the cited study, pointing out her own interpretation that the studies failed to determine whether the eradication rate was due to sequential therapy or a higher dosage of antibiotic. Her critique suggests that M.W. embodied an additional role in Paper 3 that we had not seen in our analysis of Papers 1 or 2: the role of not just a reporter of scientific information but also that of a science critic.
M.W.’s science epistemology continues to evolve during the BioEd URE, and she discusses these changes throughout Reflections 5–9. However, she most clearly articulates how the BioEd URE influenced her epistemology in her final reflection. Because the final reflection was written a semester after the experience, M.W. has had time to reflect upon her experience during the BioEd URE.
I also think that this research project has expanded my outlook on the science field because I see how there are many variables that play into science and it’s not always straightforward and black and white. Science is more than just numbers and data; you have to interpret that data and draw patterns from the articles that you read.—Reflection 10
The final sentence in this excerpt reflects the changes we see between the papers M.W. wrote before the research experience and the paper she wrote during the URE. M.W. states that science knowledge is not only data reporting, but also includes interpretation and the drawing of her own conclusions. Later in the reflection, M.W. discusses her past self and compares what she thought about science as a freshman to how she now thinks about science.
I think as a freshman, I assumed that you were not “allowed” or that it wasn’t science if I took a stance in one direction over the other. I definitely held back my opinion in the paper because I thought that it wouldn’t be right to put what I believed in the paper because it would seem too biased. Now I know that it’s okay to put your stance in a paper, as long as you can back it up with evidence while still acknowledging the limitations of your ideas. I learned that science is a lot trickier than I originally thought because you do want to present truthful information, but you can still put what you believe based on drawing real conclusions from your own research.—Reflection 10
While a first-year student, she felt that she was not supposed to take a stance in science, but she now believes that she can present beliefs as long as they are supported by evidence. We interpret “opinion” “belief” and “stance” in this excerpt as M.W.’s own conclusions drawn from the data she presents.
M.W. Develops Agency toward Scientific Knowledge Construction during the BioED URE
M.W.’s realization that science requires interpretation of data, coupled with her comments about not having room to decide what is right or wrong in her STEM class and holding back her opinion in her papers, shows that she did not feel that it was proper for her to construct knowledge in the context of a classroom. However, her critique of the research paper in Reflection 3 and the shift in her writing style in Paper 3 led us to believe M.W. developed agency toward knowledge construction during the BioED URE. We define agency as an individual’s perceived capacity to act and make choices independently within a specific structure ( Archer, 2002 ). In our case, the structure refers to constructing knowledge in the discipline of biology. However, because agency is a concept that focuses on an individual’s perceived capacity to act with intentionality ( Archer, 2002 ), it is not possible for us, as researchers outside M.W.’s mind, to draw concrete conclusions about her agency. Therefore, we explained the concept of agency to M.W. and asked her to respond to our interpretation. M.W. explains how participation in the URE affected her agency toward forming her own conclusions in her response to our analysis.
This URE taught me what agency is and how agency is valuable in the scientific world. That’s why my reflections show how I started to see how science is not just the statement and summarization of data, but the interpretation of results. This URE taught me that my ideas and my opinions matter, as long as I back up my interpretation with data, I have the ability to make my own conclusions. Although I still feel like being an undergraduate student comes with hesitation from others to accept the conclusions I make, I am confident in my ability to make those conclusions on my own. If I had not been assigned to read and reflect on the research article that I found to be misleading or be encouraged to critique articles that I read, I do not believe that I would have developed agency in my scientific writing.
Through M.W.’s response, we conclude that one of her reasons for interpreting and drawing conclusions from published data is because she feels that she has the capacity to do so. She feels that she has the agency to make independent conclusions from published data. Upon review of our analysis, M.W. wrote the following response, summarizing her views about her feelings of agency in her classes and the BioEd URE.
Having agency matters to me in determining my stance on health care because it’s a topic that is going to stick with me for the rest of my life. My understanding of STEM really only matters to the extent that I understand it enough for the test in my class. Therefore, whether or not I had agency in the context of the STEM classroom did not seem important to my learning at the time I wrote the reflection because I was just trying to earn a good grade in the course. When I read the article that I was assigned to write a reflection on, I honestly remember being annoyed with the author. The abstract was misleading; I read through the paper and felt that the abstract made a way too broad, overarching claim that I did not feel was completely supported in their research.
In her response, M.W. revisits her first reflection, commenting on how the different STEM classroom and healthcare contexts influenced her scientific agency. Forming her own conclusions was not an important goal in the STEM class, as the assessments only considered the instructor’s information as knowledge. As such, whether or not M.W. felt the agency to construct her own conclusions was moot, because her goal was non-epistemic: “to earn a good grade in the course.” She contrasts the STEM course structure with the paper critique during the BioEd URE, where she felt there was a space for her to construct her own opinion. Critiquing the paper resulted in an emotional response wherein she felt frustrated with the conclusions drawn by the authors. This emotion is important, as it can serve as motivation, in M.W.’s case, to challenge the claims of others. This experience seems to have transferred to M.W.’s writing in Paper 3, where she challenges the conclusions of one of her sources.
In this paper, we analyze one student’s biology literature reviews from three classes and written reflections to determine how she thinks about the nature of biology knowledge and its construction before and during participation in a BioEd URE. This analysis is supplemented by the student researcher, M.W., who describes her experience through an autoethnographic lens. Analysis of M.W.’s reflections and classroom papers suggests that she came to realize that she could critique knowledge produced by science experts, which led to the development of her agency toward scientific knowledge production.
Reflexivity Helped M.W. Refine Her Thoughts about Biology Knowledge Construction and Develop Scientific Agency
M.W.’s written reflections give us insight into her reflexivity, defined as the internal conversation that helps an individual to evaluate and re-evaluate their actions and decisions ( Archer, 2012 ). For example, in Reflection 4, M.W. felt that she was making things up, describing her problem-solving process as “fake chemistry,” but while re-examining her actions, realized that she solved the chemistry problem by applying prior knowledge to a new context. Through the BioEd URE and other experiences, M.W. gained an awareness about her own ability to apply concepts to challenge questions. M.W.’s examination of her own actions resulted in a change in her thinking about how she constructs solutions to problems, a hallmark of reflexivity ( Archer, 2010 ; Weinstock et al. , 2017 ).
Participating in research experiences has been shown to enhance scientific agency and project ownership ( Hester et al. , 2018 ), but less is known about how that agency develops during the experience. By making her reflexive practice explicit, M.W. helped to fill this gap by providing insight into how her scientific agency developed over the course of the BioEd URE. It is evident from M.W.’s third reflection that asking her to critique a scientific journal article was an important part of her scientific agency development. However, for her to develop scientific agency, M.W. had to first recognize how she thought about scientific knowledge and that she thought about scientific knowledge differently between contexts. In using reflexivity to examine these contexts, M.W. found that she felt little agency toward constructing knowledge in her STEM course, because in that context, the instructor decides what counts as knowledge. However, in the context of the BioEd URE, M.W. felt that her own ideas could count as knowledge, so long as she could support her ideas with evidence. We hypothesize that the structure of the training for the BioEd URE contributed to the development of M.W.’s agency toward scientific knowledge construction. Other researchers have also found differences between students’ views of knowledge within their courses and research experiences ( Faber et al. , 2016 ; Faber and Benson, 2017 ).
Possible Influences the BioEd URE Structure Had on M.W.’s Feelings of Agency toward Scientific Knowledge Construction
While M.W.’s reflections were an important part of the development of her agency, it is important to remember that the reflections were embedded within the structure we provided in the BioEd URE that was designed to help M.W. explore ways of knowing in science while embodying the role of a knowledge builder. We cannot definitively say what aspects of the BioEd URE or other educational experiences were integral for M.W.’s development of science agency. However, the development of M.W.’s feelings of agency toward scientific knowledge production could be explained through the interaction between structure and agency. Structure refers the roles that are made available to agents and the systems that maintain these roles ( Case, 2013 ), which influence the kinds of intentional actions that individuals can take ( Akram, 2013 ). The venue we provided for M.W. to share her conclusions for critique provided a role for M.W. that included agency as a fellow knowledge builder ( Longino, 2002 ). However, her conceptualization of her STEM course only provided M.W. with the role of an information gatherer. As a result, whether or not M.W. felt a sense of scientific agency was not important, because the perceived structure of the STEM class did not provide a space for M.W.’s intentional knowledge-building actions. These two examples illustrate the important role that structure plays in the development of scientific agency ( Case, 2013 ; Schenkel et al. , 2019 ).
Our results suggest that the structure that we provided during the BioEd URE played a role in the development of M.W.’s scientific agency, along with her other educational experiences. We designed our BioEd URE to ensure that the structure provided a space where M.W. could develop a feeling of scientific agency. As discussed in the overview of the URE, the design of the experience incorporated the four norms of scientific knowledge production outlined by Longino (2002) . Ensuring that M.W. felt tempered intellectual equality in the venues that we provided for critique presented M.W. with a space where she could act intentionally to construct knowledge. Furthermore, our explicit discussions about discipline-specific epistemology helped to outline the public standards of quality in the context of biology and education research, which gave M.W. the tools to evaluate her own claims.
An important part of the structure was the assignment that required M.W. to critique a published journal article. This assignment helped M.W. realize that she is allowed to critique published knowledge and that she is not required to blindly trust published information. This realization strengthened her role in science knowledge production and led to her feeling more like an agent in the production of scientific knowledge. In her response to our analysis, M.W. explicitly stated: “If I had not been assigned to read and reflect on the research article that I found to be misleading or be encouraged to critique articles that I read, I do not believe that I would have developed agency in my scientific writing.”
Another important aspect of the BioEd URE structure was the assignment of written reflections, which facilitated her reflexivity. The reflection prompts grew out of discussions in analysis meetings during the BioEd URE. For example, Reflection 3 came from a discussion about M.W.’s perception that she was not in a position to challenge the claims made by researchers. In that discussion, D.L. and D.D.-R. established the importance of M.W.’s independent analysis in the context of the BioEd URE. In doing so, D.L. and D.D.-R. established a norm for the knowledge (epistemic) culture ( Knorr-Cetina, 1999 ) of the BioEd URE. M.W. internalizes this norm in her responses to our analysis, noting that, in this URE, “my ideas and my opinions matter, as long as I back up my interpretation with data.” In Reflection 10, M.W. incorporates this epistemic norm into her EMK about science knowledge, saying: “Science is more than just numbers and data, you have to interpret that data and draw patterns from the articles that you read.” This refined idea about scientific knowledge construction helped to form M.W.’s agency toward scientific knowledge construction, because it established her role as an active agent in the interpretation of scientific data and the construction of scientific knowledge.
This paper expands on research that explores the connection between epistemic thinking and researcher identity formation in undergraduate engineering students. Much like M.W.’s experience, the work in engineering found that participants formed their ideas about knowledge generation through reflexivity. Participants compared their newly formed ideas to their own research actions and social interactions, influencing their researcher identities ( Faber et al. , 2019 ). While our paper does not explicitly ask questions about identity, the emergence of agency in our thematic analysis makes this discussion relevant, because identity is deeply interwoven with agency. An individual’s sense of self (identity) has been shown to dictate the intentional actions taken (agency) in a given context ( Archer, 2002 ). Epistemic discussions during the URE helped M.W. form her EMK about knowledge production in the context of the BioEd URE. Specifically, M.W. constructed knowledge of herself as a knowledge producer, providing a space in which she could intentionally enact the actions of a knowledge generator. These discussions support and extend previous research showing that explicit instruction on science epistemology enhances students’ understanding of the NoS ( McDonald, 2010 ; Bell et al. , 2011 ).
Study Limitations
The primary limitations associated with this study are related to the study sample, data collection, and subject as researcher. It is important to note that the study we present in this paper was developed in response to interesting insights from one student participating in a BioEd URE, and thus was not planned from the beginning as a case study with autoethnographic approaches. Because this paper describes an individual student’s experience in a URE, the results should not be generalized beyond the study context. Additionally, M.W. is a high-achieving honors student and cannot be counted as representative of an “average student.” However, the combination of case study and autoethnographic approaches facilitated the construction of an in-depth description that provides an example of how a student developed her science epistemology and scientific agency. It is also important to note that the BioEd URE was intentionally designed around epistemology. As such, results from this study cannot be generalized to biology research experiences that do not include discussions around how knowledge is generated, assessed, and justified. However, there is evidence that discussion of science epistemology in the BioEd URE influenced how M.W. approached knowledge construction in her biology course work. Therefore, we believe that biology instructors and research mentors can use the general structure of our BioEd URE as an example of how epistemic discussions can be integrated into an URE.
The data we analyzed in this study were generated by M.W. for multiple classes and were not designed specifically to answer our research questions. The conclusions we draw from these data, specifically the development of her science epistemology and her feelings of scientific agency, therefore cannot be causally connected to M.W.’s participation in the BioEd URE. In particular, M.W.’s previous research experience as well as her participation in her psychology courses and the science writing course may have significantly influenced her epistemic development. Consequently, we do not claim that the BioEd URE caused M.W. to develop science epistemology or scientific agency; instead, we attribute these developments to her whole experience as an undergraduate student. Additionally, M.W.’s involvement in the URE lasted only one semester because of curricular and time constraints. If her experience had spanned several semesters, it may have influenced her overall experience and the results of this study.
Including M.W. as a researcher who used self-analysis to bring additional insights into our work helped to address both theoretical and ethical validity; however, it also brought challenges to communicative validity and process reliability. By introducing M.W. to the theoretical concepts of epistemology and agency, we introduced the possibility that her analysis would consist of what she felt we wanted to hear as researchers. With respect to our interpretation of her epistemic development, this limitation is of less concern, as she would need to be aware of and understand her own epistemology in order to tell us what we wanted to hear. Likewise with scientific agency, we cannot be certain that her increased feelings of agency are directly associated with her new understandings of science epistemology. We (D.D.-R. and D.L.) did observe M.W. exercising her scientific agency through the BioEd URE, which allows us to begin to triangulate her responses that are associated with her experience in the BioEd URE. These limitations are not unique to this work and are shared across all studies that use self-reported data to some capacity.
With that said, before asking M.W. to be a participant researcher and as part of the BioEd URE, we discussed the quality framework described in this paper and stressed the importance of presenting authentic experience as opposed to what we wanted to hear. There is also evidence in M.W.’s research papers that suggest she developed feelings of scientific agency between writing her second and third research papers. Finally, as qualitative researchers, we are at the mercy of what our study participants are willing to share. While we stress the importance of data authenticity to our participants and triangulate our interpretations among different forms of data, in the end, we must trust what our participants share on some level. M.W. has given us no reason to doubt the authenticity of her accounts.
Implications
The reflections that M.W. wrote during the BioEd URE made explicit her thinking about scientific knowledge and may have also helped her to reify her thoughts about scientific knowledge construction. For many students, their own ways of knowing are tacit ( Hofer, 2004 ), and reflective writing could be one way for students to make explicit and evaluate these ways of knowing. Scientific writing has been found to help students develop reasoning skills in both K–12 ( Tytler and Prain, 2010 ) and higher education learning environments ( Quitadamo and Kurtz, 2007 ). During the BioEd URE, M.W. engaged in both scientific and reflective writing, which helped to activate her reflexivity, leading to development of her ideas about knowledge production. M.W.’s learning process mirrors the experiential learning cycle, in which learners reflectively observe (RO) concrete experiences (CE), helping them to construct abstract conceptualizations (AC) that can later be tested through active experimentation (AE) as other concrete experiences ( Kolb et al. , 2001 ). M.W.’s written reflections (RO) helped her to process her experiences (CE) during the URE. She also wrote about her initial thoughts about knowledge production (AC), which she could test during discussions with D.L. and D.D.-R. (AE). Our description of M.W.’s learning process has implications for teaching practice. While written reflection has been shown to enhance learning, our results suggest that once students have finished reflecting, educators should ensure that students are provided the opportunity to apply and test their abstract conceptualizations in new contexts. In this way, students will have opportunities to complete their learning cycles ( Kolb et al. , 2001 ).
An additional implication for teaching practice comes from M.W.’s responses to our analysis. D.L. and D.D.-R. interpreted a pattern of composing paragraphs primarily with paraphrased information (often with some inaccuracies) and concluding with a citation as indicating a lack of EMK of scientific knowledge construction and a lack of interpretation or synthesis of information. Based on M.W.’s input, it became clear that a lack of synthesis might actually be a lack of agency or the perception that student scientific agency is not valued in the classroom. Moreover, mistakes or misconceptions in scientific writing might actually indicate an attempt at synthesis. The challenge is for instructors to show students that constructing conclusions is valued as much as producing accurate descriptions of phenomena. Of course, biology educators do not want students conjuring false conclusions. As such, educators should provide venues for students to present their work for critique, so that students may discuss the accepted standards of science and acquire the cognitive tools necessary to produce accurate descriptions.
CONCLUSIONS
UREs provide opportunities for undergraduates to engage in the process of constructing scientific knowledge. Through this case study, we found that one student’s 1) thoughts about science epistemology differed between contexts, 2) perceptions of her role as a critic of published knowledge changed over the course of the study, and 3) feelings of agency toward knowledge construction developed during her time in the BioEd URE. While we cannot draw causal relationships between these claims and the BioEd URE, our analysis of reflections that M.W. wrote during the BioEd URE illustrate part of the reflexive process that facilitated M.W.’s epistemic development. Our work also reveals the importance of context, specifically the structure of the learning environment in the development of one student’s science epistemology and scientific agency.
Acknowledgments
This work was not supported financially by any funding agency. The authors would like to thank Elisabeth Schussler and Rachel McCord Ellestad for critical reading and comments on the manuscript.
- Akram, S. (2013). Fully unconscious and prone to habit: The characteristics of agency in the structure and agency dialectic . Journal for the Theory of Social Behaviour , 43 ( 1 ), 45–65. 10.1111/jtsb.12002 [ CrossRef ] [ Google Scholar ]
- American Association for the Advancement of Science. (2011). Vision and change in undergraduate biology education: A call to action . Washington, DC. Retrieved August 15, 2016, from https://live-visionandchange.pantheonsite.io/wp-content/uploads/2013/11/aaas-VISchange-web1113.pdf [ Google Scholar ]
- Archer, M. (2002). Realism and the problem of agency . Alethia , 5 ( 1 ), 11–20. 10.1558/aleth.v5i1.11 [ CrossRef ] [ Google Scholar ]
- Archer, M. S. (2010). Routine, reflexivity, and realism . Sociological Theory , 28 ( 3 ), 272–303. [ Google Scholar ]
- Archer, M. S. (2012). The reflective imperative in late modernity . New York, NY: Cambridge University Press. [ Google Scholar ]
- Barzilai, S., Zohar, A. (2014). Reconsidering personal epistemology as metacognition: A multifaceted approach to the analysis of epistemic thinking . Educational Psychologist , 49 ( 1 ). 10.1080/00461520.2013.863265 [ CrossRef ] [ Google Scholar ]
- Baxter, P., Jack, S. (2008). Qualitative case study methodology: Study design and implementation for novice researchers . Qualitative Report , 13 ( 4 ), 544–559. [ Google Scholar ]
- Baxter Magolda, M. B. (1992). Knowing and reasoning in college: Gender-related patterns in students’ intellectual development . San Francisco, CA: Jossey-Bass. [ Google Scholar ]
- Bell, R. L., Matkins, J. J., Gansneder, B. M. (2011). Impacts of contextual and explicit instruction on preservice elementary teachers’ understandings of the nature of science . Journal of Research in Science Teaching , 48 ( 4 ), 414–436. 10.1002/tea.20402 [ CrossRef ] [ Google Scholar ]
- Bickmore, B. R., Thompson, K. R., Grandy, D. A., Tomlin, T. (2009). Science as storytelling for teaching the nature of science and the science-religion interface . Journal of Geoscience Education , 57 ( 3 ), 178–190. 10.5408/1.3544263 [ CrossRef ] [ Google Scholar ]
- Case, J. M. (2013). Researching student learning in higher education: A social realist approach . New York, NY: Routledge. [ Google Scholar ]
- Chinn, C., Rinehart, R., Buckland, A. (2014). Epistemic cognition and evaluating information: Applying the AIR model of epistemic cognition . In Rapp, D., Braasch, J. (Eds.), Processing inaccurate information: Theoretical and applied perspectives from cognitive science and the educational sciences (pp. 425–453). Cambridge, MA: MIT Press. [ Google Scholar ]
- Creswell, J. (2012). Qualitative research & design: Choosing among five approaches (3rd ed.). Thousand Oaks, CA: Sage. [ Google Scholar ]
- Elby, A., Hammer, D. (2001). On the substance of a sophisticated epistemology . Science Education , 85 ( 5 ), 554–567. [ Google Scholar ]
- Ellis, C., Adams, T. E., Bochner, A. P. (2011). Autoethnography: An overview . Historical Social Research , 36 ( 4 ), 273–290. [ Google Scholar ]
- Faber, C., Benson, L. C. (2017). Engineering students’ epistemic cognition in the context of problem solving: epistemic cognition in problem solving . Journal of Engineering Education , 106 ( 4 ), 677–709. 10.1002/jee.20183 [ CrossRef ] [ Google Scholar ]
- Faber, C., Vargas, P., Benson, L. (2016). Engineering students’ epistemic cognition in a research environment . International Journal of Engineering Education , 32 ( 16 ) [ Google Scholar ]
- Faber, C. J., Benson, L. C., Kajfez, R. L., Kennedy, M. S., Lee, D. M., McAlister, A. M., ... & Wu, G. (2019). Dynamics of researcher identity and epistemology: The development of a grounded-theory model . Paper presented at: American Society for Engineering Education Conference held from June 15–19, 2019 (Tampa, FL) .
- Gottesman, A. J., Hoskins, S. G. (2013). CREATE cornerstone: Introduction to scientific thinking, a new course for STEM-interested freshmen, demystifies scientific thinking through analysis of scientific literature . CBE—Life Sciences Education , 12 ( 1 ), 59–72. 10.1187/cbe.12-11-0201 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Hammer, D., Elby, A., Scherr, R. E., Redish, E. F., Hammer, D., Elby, A., ... & Redish, E. F. (2005). Resources, framing, and transfer . In Mestre, J. (Ed.), Transfer of learning from a modern multidisciplinary perspective (pp. 89–120). Greenwich, CT: Information Age. [ Google Scholar ]
- Hester, S. D., Nadler, M., Katcher, J., Elfring, L. K., Dykstra, E., Rezende, L. F., Bolger, M. S. (2018). Authentic Inquiry through Modeling in Biology (AIM-Bio): An introductory laboratory curriculum that increases undergraduates’ scientific agency and skills . CBE—Life Sciences Education , 17 ( 4 ), ar63. 10.1187/cbe.18-06-0090 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Hofer, B. K. (2004). Epistemological understanding as a metacognitive process: Thinking aloud during online searching . Educational Psychologist , 39 ( 1 ), 43–55. 10.1207/s15326985ep3901_5 [ CrossRef ] [ Google Scholar ]
- Hofer, B. K., Pintrich, P. R. (1997). The development of epistemological theories: Beliefs about knowledge and knowing and their relation to learning . Review of Educational Research , 67 ( 1 ), 88–140. 10.3102/00346543067001088 [ CrossRef ] [ Google Scholar ]
- Hoskins, S. G., Lopatto, D., Stevens, L. M. (2011). The C.R.E.A.T.E. approach to primary literature shifts undergraduates’ self-assessed ability to read and analyze journal articles, attitudes about science, and epistemological beliefs . CBE—Life Sciences Education , 10 ( 4 ), 368–378. 10.1187/cbe.11-03-0027 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Hughes, S., Pennington, J. L., Makris, S. (2012). Translating autoethnography across the AERA standards: Toward understanding autoethnographic scholarship as empirical research . Educational Researcher , 41 ( 6 ), 209–219. 10.3102/0013189X12442983 [ CrossRef ] [ Google Scholar ]
- Hunter, A., Laursen, S. L., Seymour, E. (2007). Becoming a scientist: The role of undergraduate research in students’ cognitive, personal, and professional development . Science Education , 91 ( 1 ), 36–74. 10.1002/sce [ CrossRef ] [ Google Scholar ]
- Jimenez-Aleixandre, M. P. J., Rodriguez, A. B., Duschl, R. A. (1999). “Doing the lesson” or “doing science”: Argument in high school genetics Science education , 84 ( 6 ), 757–792. [ Google Scholar ]
- Kalman, C. S. (2007). Successful science and engineering teaching in colleges and universities . Bolton, MA: Anker Publishing. [ Google Scholar ]
- Kelly, G. J. (2008). Inquiry, activity and epistemic practice . In Duschl, R., Grandy, R. (Eds.), Teaching scientific inquiry: Recommendations for research and implementation (pp. 99–117). Rotterdam: Sense Publishers. [ Google Scholar ]
- Kenyon, K. L., Onorato, M. E., Gottesman, A. J., Hoque, J., Hoskins, S. G. (2016). Testing CREATE at community colleges: An examination of faculty perspectives and diverse student gains . CBE—Life Sciences Education , 15 ( 1 ), 1–19. 10.1187/cbe.15-07-0146 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- King, P. M., Kitchener, K. S. (1994). Developing reflective judgment: Understanding and promoting intellectual growth and critical thinking in adolescents . San Francisco, CA: Jossey-Bass. [ Google Scholar ]
- Knorr-Cetina, K. (1999). Epistemic cultures: How the sciences make knowledge . Cambridge, MA: Harvard University Press. [ Google Scholar ]
- Kolb, D. A., Boyatzis, R. E., Mainemelis, C. (2001). Experiential learning theory: Previous research and future directions . In Sternberg, R. J., Zhang, L.-F. (Eds.), Perspectives on thinking, learning and cognitive styles (pp. 227–247). Mahwah, NJ: Erlbaum. [ Google Scholar ]
- Kuhn, D. (1991). The skills of argument . New York, NY: Cambridge University Press. [ Google Scholar ]
- Lederman, N. G. (2007). Nature of science: Past, present, and future . In Abell, S. K.., Lederman, N. G. (Eds.), Handbook of research on science education . New York, NY: Routledge. [ Google Scholar ]
- Lee, D. M., McAlister, A. M., Ehlert, K., Faber, C. J., Kajfez, R. L., Creamer, E., Kennedy, M. S. (2019). The use of analytical memo writing in a mixed methods grounded theory study . Paper presented at: Frontiers in Education (FIE) Conference held from October 16–19, 2019 (Cincinnati, OH) .
- Linn, M. C., Palmer, E., Baranger, A., Gerard, E., Stone, E. (2015). Undergraduate research experiences: Impacts and opportunities . Science , 347 ( 6222 ), 1261757–1261757. 10.1126/science.1261757 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Longino, H. E. (2002). The fate of knowledge . Princeton, NJ: Princeton University Press. [ Google Scholar ]
- Lopatto, D. (2004). Survey of undergraduate research experiences (SURE): First findings . Cell Biology Education , 3 ( 4 ), 270–277. 10.1187/cbe.04-07-0045 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Lopatto, D. (2007). Undergraduate research experiences support science . CBE—Life Sciences Education , 6 , 297–306. 10.1187/cbe.07 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Louca, L., Elby, A., Hammer, D., Kagey, T. (2004). Epistemological resources: Applying a new epistemological framework to science instruction . Educational Psychologist , 39 ( 1 ), 57–68. 10.1207/s15326985ep3901_6 [ CrossRef ] [ Google Scholar ]
- McDonald, C. V. (2010). The influence of explicit nature of science and argumentation instruction on preservice primary teachers’ views of nature of science . Journal of Research in Science Teaching , 47 ( 9 ), 1137–1164. 10.1002/tea.20377 [ CrossRef ] [ Google Scholar ]
- National Academies of Sciences, Engineering, and Medicine. (2017). Undergraduate research experiences for STEM students: Successes, challenges, and opportunities (pp. 24622). Washington, DC: National Academies Press. 10.17226/24622 [ CrossRef ] [ Google Scholar ]
- National Research Council (NRC). (2007). Taking science to school: Learning and teaching science in grades K–8 . Washington, DC: National Academies Press. [ Google Scholar ]
- NRC. (2013). Next Generation Science Standards: For states, by states . Washington, DC: National Academies Press. [ Google Scholar ]
- Perry, W. G. (1990). Forms of ethical and intellectual development in the college years: A scheme . San Francisco, CA: Jossey-Bass. [ Google Scholar ]
- Polkinghorne, D. E. (1995). Narrative configuration in qualitative analysis . International Journal of Qualitative Studies in Education , 8 ( 1 ), 5–23. [ Google Scholar ]
- Quitadamo, I. J., Kurtz, M. J. (2007). Learning to improve: Using writing to increase critical thinking performance in general education biology . CBE—Life Sciences Education , 6 , 140–154. 10.1187/cbe.06 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Ryu, S., Sandoval, W. A. (2012). Improvements to elementary children’s epistemic understanding from sustained argumentation . Science Education , 96 ( 3 ), 488–526. 10.1002/sce.21006 [ CrossRef ] [ Google Scholar ]
- Sadler, T. D., Burgin, S., McKinney, L., Ponjuan, L. (2010). Learning science through research apprenticeships: A critical review of the literature . Journal of Research in Science Teaching: The Official Journal of the National Association for Research in Science Teaching , 47 ( 3 ), 235–256. [ Google Scholar ]
- Sandoval, W. A. (2005). Understanding students’ practical epistemologies and their influence on learning through inquiry . Science Education , 89 ( 4 ), 634–656. 10.1002/sce.20065 [ CrossRef ] [ Google Scholar ]
- Sandoval, W. A. (2012). Situating epistemological development . In van Aalst, J., Thompson, K., Jacobson, J., Reimann, P. (Eds.), The future of learning: Proceedings of the 10th international conference of learning sciences (pp. 347–354). Sydney, Australia: International Society of the Learning Sciences. [ Google Scholar ]
- Schenkel, K., Calabrese Barton, A., Tan, E., Nazar, C. R., Flores, M. D. G. D. (2019). Framing equity through a closer examination of critical science agency . Cultural Studies of Science Education , 14 ( 2 ), 309–325. 10.1007/s11422-019-09914-1 [ CrossRef ] [ Google Scholar ]
- Schommer-Aikins, M., Duell, O. K., Hutter, R. (2005). Epistemological beliefs, mathematical problem-solving beliefs, and academic performance of middle school students . Elementary School Journal , 105 ( 3 ), 289–304. 10.1086/428745 [ CrossRef ] [ Google Scholar ]
- Semsar, K., Knight, J. K., Birol, G., Smith, M. K. (2011). The Colorado Learning Attitudes about Science Survey (CLASS) for use in biology . CBE—Life Sciences Education , 10 ( 3 ), 268–278. 10.1187/cbe.10-10-0133 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Seymour, E., Hunter, A.-B., Laursen, S. L., DeAntoni, T. (2004). Establishing the benefits of research experiences for undergraduate in the sciences: First findings from a three-year study . Science Education , 88 ( 4 ), 493–534. 10.1002/sce.10131 [ CrossRef ] [ Google Scholar ]
- Sochacka, N. W., Walther, J., Pawley, A. L. (2018). Ethical validation: Reframing research ethics in engineering education research to improve research quality . Journal of Engineering Education , 107 ( 3 ), 362–379. 10.1002/jee.20222 [ CrossRef ] [ Google Scholar ]
- Stake, R. E. (2000). Case studies . In Denzin, N. K., Lincoln, Y. S. (Eds.), Handbook of qualitative research (2nd ed., pp. 435–454). Thousand Oaks, CA: Sage. [ Google Scholar ]
- Stake, R. E. (2006). Multiple case study analysis . New York, NY: Guilford. [ Google Scholar ]
- Stanton, J. D., Dye, K. M., Johnson, M. (2019). Knowledge of learning makes a difference: A comparison of metacognition in introductory and senior-level biology students . CBE—Life Sciences Education , 18 ( 2 ), ar24. 10.1187/cbe.18-12-0239 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Tan, E., Barton, A. C. (2008a). From peripheral to central, the story of Melanie’s metamorphosis in an urban middle school science class . Science Education , 92 ( 4 ), 567–590. 10.1002/sce.20253 [ CrossRef ] [ Google Scholar ]
- Tan, E., Barton, A. C. (2008b). Unpacking science for all through the lens of identities-in-practice: The stories of Amelia and Ginny . Cultural Studies of Science Education , 3 , 43–71. [ Google Scholar ]
- Thiry, H., Laursen, S. L., Hunter, A. B. (2005). What experiences help students become scientists? A comparative study of research and other sources of personal and professional gains for STEM undergraduates . The Journal of Higher Education , 82 ( 4 ), 357–388. [ Google Scholar ]
- Thiry, H., Weston, T., Laursen, S., Hunter, A. (2012). The benefits of multi-year research experiences: Differences in novice and experienced students’ reported gains from undergraduate research . CBE—Life Sciences Education , 11 ( 3 ), 260–272. 10.1187/cbe.11-11-0098 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Toulmin, S. E. (2003). The uses of argument, Updated edition (2nd ed.). New York, NY: Cambridge University Press. [ Google Scholar ]
- Tytler, R., Prain, V. (2010). A framework for re-thinking learning in science from recent cognitive science perspectives . International Journal of Science Education , 32 ( 15 ), 2055–2078. 10.1080/09500690903334849 [ CrossRef ] [ Google Scholar ]
- Venville, G., Gribble, S. J., Donovan, J. (2005). An exploration of young children’s understandings of genetics concepts from ontological and epistemological perspectives . Science Education , 89 ( 4 ), 614–633. 10.1002/sce.20061 [ CrossRef ] [ Google Scholar ]
- Walker, J. D., Cotner, S. H., Baepler, P. M., Decker, M. D. (2008). A delicate balance: Integrating active learning into a large lecture course . CBE—Life Sciences Education , 7 , 361–367. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Walther, J., Sochacka, N. W., Kellam, N. N. (2013). Quality in interpretive engineering education research: Reflections on an example study . Journal of Engineering Education , 102 ( 4 ), 626–659. 10.1002/jee.20029 [ CrossRef ] [ Google Scholar ]
- Watkins, J., Elby, A. (2013). Context dependence of students’ views about the role of equations in understanding biology . CBE—Life Sciences Education , 12 ( 2 ), 274–286. 10.1187/cbe.12-11-0185 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Weinstock, M., Kienhues, D., Feucht, F. C., Ryan, M. (2017). Informed reflexivity: Enacting epistemic virtue . Educational Psychologist , 52 ( 4 ), 284–298. 10.1080/00461520.2017.1349662 [ CrossRef ] [ Google Scholar ]
- Yin, R. K. (2018). Case study research and applications: Design and methods (6th ed.). Thousand Oaks, CA: Sage. [ Google Scholar ]

The Official Blog of Edvotek®
The Biotechnology Education Company | Kits & Equipment Designed for the Classroom Since 1987

Qualitative vs. Quantitative Research in Biotechnology

If you’ve read (or taught) a biology textbook you’ve probably come across a section on qualitative and quantitative data. Chances are it was brief with basic definitions and maybe a mention that that both types of data are important to science. But if you’re here on our blog you’ve likely leaped a more hands-on, experiment-centric curriculum and are looking for a deeper dive into the two q’s of data. Read on!
Qualitative and quantitative describe distinct types of data as well as slightly different approaches to research. Here’s how they differ:
- Qualitative research focuses on non-numeric data. It involves collecting and analyzing descriptive, narrative, or textual information, often in the form of words, images, or observations. Qualitative data can be open-ended, subjective, and rich in detail.
- Quantitative research deals with numeric data. It involves collecting data in the form of numbers, such as measurements, counts, or ratings. The data is typically analyzed using statistical techniques. A consequence of this is that quantitative research often involves larger sample sizes and data sets to ensure statistical validity.
The Methods
- Qualitative research methods include observations as well as case studies, interviews, and focus groups. Researchers seek to understand and interpret underlying meanings, patterns, and contexts. Researchers use themes, patterns, and narrative explanations to make sense of the data.
- Quantitative research methods include experiments, structured observations, surveys, and data analysis through statistical tools and techniques. Researchers aim to quantify relationships and trends in data. Analysis in quantitative research involves statistical tests and calculations to assess relationships, significance, and patterns within the numerical data. Researchers use themes, patterns, and narrative explanations to make sense of the data but these findings are supported by calculated values like means and correlation coefficients.
The Objectives
- Qualitative research is exploratory. It’s often used to generate hypotheses, uncover new insights, and develop a narrative. Qualitative data in medical diagnosis involves the subjective assessment of symptoms, patient narratives, and contextual information. Some diagnosis tests are also qualitative or a hybrid of the two methods (semi-quantitative).
- Quantitative research is used to test hypotheses, establish relationships, and make predictions. It focuses on measuring variables, determining cause-and-effect relationships, and generalizing findings for large data sets. Quantitative data in medical diagnosis involves the numerical analysis of measurable parameters, such as laboratory values or imaging results, providing objective and standardized information to support clinical assessments.
Researchers choose between qualitative and quantitative methods based on their research questions, objectives, and the nature of the data they want to collect and analyze. In some cases, a mixed-methods approach, combining both qualitative and quantitative methods, is used to provide a more comprehensive understanding of a research topic.
In biotechnology, both types of data are often used together. Qualitative data may help researchers identify interesting phenomena or patterns, while quantitative data allows for precise measurement and statistical analysis. For example, qualitative observations of abnormal cell morphology might lead to quantitative measurements of cell counts or the concentration of a specific molecule to understand the underlying causes of the observed changes. The combination of qualitative and quantitative data is a powerful approach to biotechnology research and applications.
Examples of Qualitative Research in Biotechnology
Qualitative data in biotechnology often involve descriptive observations of biological processes. This can include visual assessments of cell morphology, tissue structure, or the appearance of microorganisms. Qualitative data can also include the presence or absence of specific molecules or genes in a biological sample (although depending on how this presence/absence data is processed it can also be quantitative). Qualitative data may involve the assessment of phenotypic characteristics like color, shape, growth patterns, or other observable traits in organisms or cell cultures. Microscopy and staining is a common method in biotechnology to obtain qualitative data. Researchers can use various staining techniques to visualize cellular structures, pathogens, or cellular localization of specific molecules. Techniques like PCR (Polymerase Chain Reaction) and ELISA (Enzyme Linked Immunosorbent Assay) can be used to detect the presence of specific DNA sequences or molecules.
Examples of Quantitative Research in Biotechnology
Quantitative data in biotechnology involves measuring biological variables using numeric values. For instance, researchers may measure the concentration of specific molecules or sequences or quantify the number of cells in a culture. Popular biotechnologies that enable quantitative research are: Spectrophotometric analysis (used to quantitatively determine the concentration of substances by measuring the absorption of light at specific wavelengths), Real-time PCR (used to quantitatively measure the amount of DNA or RNA in a sample), ELISAs with a standard curve (use to measure the concentration of certain molecules) and Flow Cytometry (used to measures the characteristics of cells or particles in a fluid stream, such as cell size, granularity, and the presence of specific surface markers).
Last, but not least, help to remember the difference!
Qualitative starts with “Qual,” and you can associate it with “Quality.” Think of qualitative data as focusing on the quality of information, emphasizing descriptive details, and the “how” and “why” of a phenomenon. Quantitative starts with “Quant” and you can associate it with “Quantity” (the amount of something) or even just the presets of a “N” for numbers. Or you can try this little rhyme or any part thereof:
Qualitative is tales and tales, while Quantitative counts and scales.
Qualitative, tales we create, in colors and details, it’s truly great. Quantitative, counts on a scale, with numbers precise, it’ll never fail.
Qualitative is a colorful show, it tells the ‘what’ with a creative flow. Quantitative, numbers align, it counts the ‘how much,’ it’s data’s goldmine.
In Qualitative, details bloom, painting pictures that chase away the gloom. Quantitative, numbers at play, measuring data in a structured array.
Check out some of our popular qualitative and quantitative experiments for the classroom.
- Click to share on Twitter (Opens in new window)
- Click to share on Facebook (Opens in new window)
- Already have a WordPress.com account? Log in now.
- Subscribe Subscribed
- Copy shortlink
- Report this content
- View post in Reader
- Manage subscriptions
- Collapse this bar
Have a language expert improve your writing
Run a free plagiarism check in 10 minutes, generate accurate citations for free.
- Knowledge Base
Methodology
- What Is Qualitative Research? | Methods & Examples
What Is Qualitative Research? | Methods & Examples
Published on June 19, 2020 by Pritha Bhandari . Revised on June 22, 2023.
Qualitative research involves collecting and analyzing non-numerical data (e.g., text, video, or audio) to understand concepts, opinions, or experiences. It can be used to gather in-depth insights into a problem or generate new ideas for research.
Qualitative research is the opposite of quantitative research , which involves collecting and analyzing numerical data for statistical analysis.
Qualitative research is commonly used in the humanities and social sciences, in subjects such as anthropology, sociology, education, health sciences, history, etc.
- How does social media shape body image in teenagers?
- How do children and adults interpret healthy eating in the UK?
- What factors influence employee retention in a large organization?
- How is anxiety experienced around the world?
- How can teachers integrate social issues into science curriculums?
Table of contents
Approaches to qualitative research, qualitative research methods, qualitative data analysis, advantages of qualitative research, disadvantages of qualitative research, other interesting articles, frequently asked questions about qualitative research.
Qualitative research is used to understand how people experience the world. While there are many approaches to qualitative research, they tend to be flexible and focus on retaining rich meaning when interpreting data.
Common approaches include grounded theory, ethnography , action research , phenomenological research, and narrative research. They share some similarities, but emphasize different aims and perspectives.
| Approach | What does it involve? |
|---|---|
| Grounded theory | Researchers collect rich data on a topic of interest and develop theories . |
| Researchers immerse themselves in groups or organizations to understand their cultures. | |
| Action research | Researchers and participants collaboratively link theory to practice to drive social change. |
| Phenomenological research | Researchers investigate a phenomenon or event by describing and interpreting participants’ lived experiences. |
| Narrative research | Researchers examine how stories are told to understand how participants perceive and make sense of their experiences. |
Note that qualitative research is at risk for certain research biases including the Hawthorne effect , observer bias , recall bias , and social desirability bias . While not always totally avoidable, awareness of potential biases as you collect and analyze your data can prevent them from impacting your work too much.
Receive feedback on language, structure, and formatting
Professional editors proofread and edit your paper by focusing on:
- Academic style
- Vague sentences
- Style consistency
See an example

Each of the research approaches involve using one or more data collection methods . These are some of the most common qualitative methods:
- Observations: recording what you have seen, heard, or encountered in detailed field notes.
- Interviews: personally asking people questions in one-on-one conversations.
- Focus groups: asking questions and generating discussion among a group of people.
- Surveys : distributing questionnaires with open-ended questions.
- Secondary research: collecting existing data in the form of texts, images, audio or video recordings, etc.
- You take field notes with observations and reflect on your own experiences of the company culture.
- You distribute open-ended surveys to employees across all the company’s offices by email to find out if the culture varies across locations.
- You conduct in-depth interviews with employees in your office to learn about their experiences and perspectives in greater detail.
Qualitative researchers often consider themselves “instruments” in research because all observations, interpretations and analyses are filtered through their own personal lens.
For this reason, when writing up your methodology for qualitative research, it’s important to reflect on your approach and to thoroughly explain the choices you made in collecting and analyzing the data.
Qualitative data can take the form of texts, photos, videos and audio. For example, you might be working with interview transcripts, survey responses, fieldnotes, or recordings from natural settings.
Most types of qualitative data analysis share the same five steps:
- Prepare and organize your data. This may mean transcribing interviews or typing up fieldnotes.
- Review and explore your data. Examine the data for patterns or repeated ideas that emerge.
- Develop a data coding system. Based on your initial ideas, establish a set of codes that you can apply to categorize your data.
- Assign codes to the data. For example, in qualitative survey analysis, this may mean going through each participant’s responses and tagging them with codes in a spreadsheet. As you go through your data, you can create new codes to add to your system if necessary.
- Identify recurring themes. Link codes together into cohesive, overarching themes.
There are several specific approaches to analyzing qualitative data. Although these methods share similar processes, they emphasize different concepts.
| Approach | When to use | Example |
|---|---|---|
| To describe and categorize common words, phrases, and ideas in qualitative data. | A market researcher could perform content analysis to find out what kind of language is used in descriptions of therapeutic apps. | |
| To identify and interpret patterns and themes in qualitative data. | A psychologist could apply thematic analysis to travel blogs to explore how tourism shapes self-identity. | |
| To examine the content, structure, and design of texts. | A media researcher could use textual analysis to understand how news coverage of celebrities has changed in the past decade. | |
| To study communication and how language is used to achieve effects in specific contexts. | A political scientist could use discourse analysis to study how politicians generate trust in election campaigns. |
Qualitative research often tries to preserve the voice and perspective of participants and can be adjusted as new research questions arise. Qualitative research is good for:
- Flexibility
The data collection and analysis process can be adapted as new ideas or patterns emerge. They are not rigidly decided beforehand.
- Natural settings
Data collection occurs in real-world contexts or in naturalistic ways.
- Meaningful insights
Detailed descriptions of people’s experiences, feelings and perceptions can be used in designing, testing or improving systems or products.
- Generation of new ideas
Open-ended responses mean that researchers can uncover novel problems or opportunities that they wouldn’t have thought of otherwise.
Researchers must consider practical and theoretical limitations in analyzing and interpreting their data. Qualitative research suffers from:
- Unreliability
The real-world setting often makes qualitative research unreliable because of uncontrolled factors that affect the data.
- Subjectivity
Due to the researcher’s primary role in analyzing and interpreting data, qualitative research cannot be replicated . The researcher decides what is important and what is irrelevant in data analysis, so interpretations of the same data can vary greatly.
- Limited generalizability
Small samples are often used to gather detailed data about specific contexts. Despite rigorous analysis procedures, it is difficult to draw generalizable conclusions because the data may be biased and unrepresentative of the wider population .
- Labor-intensive
Although software can be used to manage and record large amounts of text, data analysis often has to be checked or performed manually.
If you want to know more about statistics , methodology , or research bias , make sure to check out some of our other articles with explanations and examples.
- Chi square goodness of fit test
- Degrees of freedom
- Null hypothesis
- Discourse analysis
- Control groups
- Mixed methods research
- Non-probability sampling
- Quantitative research
- Inclusion and exclusion criteria
Research bias
- Rosenthal effect
- Implicit bias
- Cognitive bias
- Selection bias
- Negativity bias
- Status quo bias
Quantitative research deals with numbers and statistics, while qualitative research deals with words and meanings.
Quantitative methods allow you to systematically measure variables and test hypotheses . Qualitative methods allow you to explore concepts and experiences in more detail.
There are five common approaches to qualitative research :
- Grounded theory involves collecting data in order to develop new theories.
- Ethnography involves immersing yourself in a group or organization to understand its culture.
- Narrative research involves interpreting stories to understand how people make sense of their experiences and perceptions.
- Phenomenological research involves investigating phenomena through people’s lived experiences.
- Action research links theory and practice in several cycles to drive innovative changes.
Data collection is the systematic process by which observations or measurements are gathered in research. It is used in many different contexts by academics, governments, businesses, and other organizations.
There are various approaches to qualitative data analysis , but they all share five steps in common:
- Prepare and organize your data.
- Review and explore your data.
- Develop a data coding system.
- Assign codes to the data.
- Identify recurring themes.
The specifics of each step depend on the focus of the analysis. Some common approaches include textual analysis , thematic analysis , and discourse analysis .
Cite this Scribbr article
If you want to cite this source, you can copy and paste the citation or click the “Cite this Scribbr article” button to automatically add the citation to our free Citation Generator.
Bhandari, P. (2023, June 22). What Is Qualitative Research? | Methods & Examples. Scribbr. Retrieved August 7, 2024, from https://www.scribbr.com/methodology/qualitative-research/
Is this article helpful?

Pritha Bhandari
Other students also liked, qualitative vs. quantitative research | differences, examples & methods, how to do thematic analysis | step-by-step guide & examples, "i thought ai proofreading was useless but..".
I've been using Scribbr for years now and I know it's a service that won't disappoint. It does a good job spotting mistakes”
- Bipolar Disorder
- Therapy Center
- When To See a Therapist
- Types of Therapy
- Best Online Therapy
- Best Couples Therapy
- Best Family Therapy
- Managing Stress
- Sleep and Dreaming
- Understanding Emotions
- Self-Improvement
- Healthy Relationships
- Student Resources
- Personality Types
- Sweepstakes
- Guided Meditations
- Verywell Mind Insights
- 2024 Verywell Mind 25
- Mental Health in the Classroom
- Editorial Process
- Meet Our Review Board
- Crisis Support
Quantitative vs. Qualitative Research in Psychology
- Key Differences
Quantitative Research Methods
Qualitative research methods.
- How They Relate
In psychology and other social sciences, researchers are faced with an unresolved question: Can we measure concepts like love or racism the same way we can measure temperature or the weight of a star? Social phenomena—things that happen because of and through human behavior—are especially difficult to grasp with typical scientific models.
At a Glance
Psychologists rely on quantitative and quantitative research to better understand human thought and behavior.
- Qualitative research involves collecting and evaluating non-numerical data in order to understand concepts or subjective opinions.
- Quantitative research involves collecting and evaluating numerical data.
This article discusses what qualitative and quantitative research are, how they are different, and how they are used in psychology research.
Qualitative Research vs. Quantitative Research
In order to understand qualitative and quantitative psychology research, it can be helpful to look at the methods that are used and when each type is most appropriate.
Psychologists rely on a few methods to measure behavior, attitudes, and feelings. These include:
- Self-reports , like surveys or questionnaires
- Observation (often used in experiments or fieldwork)
- Implicit attitude tests that measure timing in responding to prompts
Most of these are quantitative methods. The result is a number that can be used to assess differences between groups.
However, most of these methods are static, inflexible (you can't change a question because a participant doesn't understand it), and provide a "what" answer rather than a "why" answer.
Sometimes, researchers are more interested in the "why" and the "how." That's where qualitative methods come in.
Qualitative research is about speaking to people directly and hearing their words. It is grounded in the philosophy that the social world is ultimately unmeasurable, that no measure is truly ever "objective," and that how humans make meaning is just as important as how much they score on a standardized test.
Used to develop theories
Takes a broad, complex approach
Answers "why" and "how" questions
Explores patterns and themes
Used to test theories
Takes a narrow, specific approach
Answers "what" questions
Explores statistical relationships
Quantitative methods have existed ever since people have been able to count things. But it is only with the positivist philosophy of Auguste Comte (which maintains that factual knowledge obtained by observation is trustworthy) that it became a "scientific method."
The scientific method follows this general process. A researcher must:
- Generate a theory or hypothesis (i.e., predict what might happen in an experiment) and determine the variables needed to answer their question
- Develop instruments to measure the phenomenon (such as a survey, a thermometer, etc.)
- Develop experiments to manipulate the variables
- Collect empirical (measured) data
- Analyze data
Quantitative methods are about measuring phenomena, not explaining them.
Quantitative research compares two groups of people. There are all sorts of variables you could measure, and many kinds of experiments to run using quantitative methods.
These comparisons are generally explained using graphs, pie charts, and other visual representations that give the researcher a sense of how the various data points relate to one another.
Basic Assumptions
Quantitative methods assume:
- That the world is measurable
- That humans can observe objectively
- That we can know things for certain about the world from observation
In some fields, these assumptions hold true. Whether you measure the size of the sun 2000 years ago or now, it will always be the same. But when it comes to human behavior, it is not so simple.
As decades of cultural and social research have shown, people behave differently (and even think differently) based on historical context, cultural context, social context, and even identity-based contexts like gender , social class, or sexual orientation .
Therefore, quantitative methods applied to human behavior (as used in psychology and some areas of sociology) should always be rooted in their particular context. In other words: there are no, or very few, human universals.
Statistical information is the primary form of quantitative data used in human and social quantitative research. Statistics provide lots of information about tendencies across large groups of people, but they can never describe every case or every experience. In other words, there are always outliers.
Correlation and Causation
A basic principle of statistics is that correlation is not causation. Researchers can only claim a cause-and-effect relationship under certain conditions:
- The study was a true experiment.
- The independent variable can be manipulated (for example, researchers cannot manipulate gender, but they can change the primer a study subject sees, such as a picture of nature or of a building).
- The dependent variable can be measured through a ratio or a scale.
So when you read a report that "gender was linked to" something (like a behavior or an attitude), remember that gender is NOT a cause of the behavior or attitude. There is an apparent relationship, but the true cause of the difference is hidden.
Pitfalls of Quantitative Research
Quantitative methods are one way to approach the measurement and understanding of human and social phenomena. But what's missing from this picture?
As noted above, statistics do not tell us about personal, individual experiences and meanings. While surveys can give a general idea, respondents have to choose between only a few responses. This can make it difficult to understand the subtleties of different experiences.
Quantitative methods can be helpful when making objective comparisons between groups or when looking for relationships between variables. They can be analyzed statistically, which can be helpful when looking for patterns and relationships.
Qualitative data are not made out of numbers but rather of descriptions, metaphors, symbols, quotes, analysis, concepts, and characteristics. This approach uses interviews, written texts, art, photos, and other materials to make sense of human experiences and to understand what these experiences mean to people.
While quantitative methods ask "what" and "how much," qualitative methods ask "why" and "how."
Qualitative methods are about describing and analyzing phenomena from a human perspective. There are many different philosophical views on qualitative methods, but in general, they agree that some questions are too complex or impossible to answer with standardized instruments.
These methods also accept that it is impossible to be completely objective in observing phenomena. Researchers have their own thoughts, attitudes, experiences, and beliefs, and these always color how people interpret results.
Qualitative Approaches
There are many different approaches to qualitative research, with their own philosophical bases. Different approaches are best for different kinds of projects. For example:
- Case studies and narrative studies are best for single individuals. These involve studying every aspect of a person's life in great depth.
- Phenomenology aims to explain experiences. This type of work aims to describe and explore different events as they are consciously and subjectively experienced.
- Grounded theory develops models and describes processes. This approach allows researchers to construct a theory based on data that is collected, analyzed, and compared to reach new discoveries.
- Ethnography describes cultural groups. In this approach, researchers immerse themselves in a community or group in order to observe behavior.
Qualitative researchers must be aware of several different methods and know each thoroughly enough to produce valuable research.
Some researchers specialize in a single method, but others specialize in a topic or content area and use many different methods to explore the topic, providing different information and a variety of points of view.
There is not a single model or method that can be used for every qualitative project. Depending on the research question, the people participating, and the kind of information they want to produce, researchers will choose the appropriate approach.
Interpretation
Qualitative research does not look into causal relationships between variables, but rather into themes, values, interpretations, and meanings. As a rule, then, qualitative research is not generalizable (cannot be applied to people outside the research participants).
The insights gained from qualitative research can extend to other groups with proper attention to specific historical and social contexts.
Relationship Between Qualitative and Quantitative Research
It might sound like quantitative and qualitative research do not play well together. They have different philosophies, different data, and different outputs. However, this could not be further from the truth.
These two general methods complement each other. By using both, researchers can gain a fuller, more comprehensive understanding of a phenomenon.
For example, a psychologist wanting to develop a new survey instrument about sexuality might and ask a few dozen people questions about their sexual experiences (this is qualitative research). This gives the researcher some information to begin developing questions for their survey (which is a quantitative method).
After the survey, the same or other researchers might want to dig deeper into issues brought up by its data. Follow-up questions like "how does it feel when...?" or "what does this mean to you?" or "how did you experience this?" can only be answered by qualitative research.
By using both quantitative and qualitative data, researchers have a more holistic, well-rounded understanding of a particular topic or phenomenon.
Qualitative and quantitative methods both play an important role in psychology. Where quantitative methods can help answer questions about what is happening in a group and to what degree, qualitative methods can dig deeper into the reasons behind why it is happening. By using both strategies, psychology researchers can learn more about human thought and behavior.
Gough B, Madill A. Subjectivity in psychological science: From problem to prospect . Psychol Methods . 2012;17(3):374-384. doi:10.1037/a0029313
Pearce T. “Science organized”: Positivism and the metaphysical club, 1865–1875 . J Hist Ideas . 2015;76(3):441-465.
Adams G. Context in person, person in context: A cultural psychology approach to social-personality psychology . In: Deaux K, Snyder M, eds. The Oxford Handbook of Personality and Social Psychology . Oxford University Press; 2012:182-208.
Brady HE. Causation and explanation in social science . In: Goodin RE, ed. The Oxford Handbook of Political Science. Oxford University Press; 2011. doi:10.1093/oxfordhb/9780199604456.013.0049
Chun Tie Y, Birks M, Francis K. Grounded theory research: A design framework for novice researchers . SAGE Open Med . 2019;7:2050312118822927. doi:10.1177/2050312118822927
Reeves S, Peller J, Goldman J, Kitto S. Ethnography in qualitative educational research: AMEE Guide No. 80 . Medical Teacher . 2013;35(8):e1365-e1379. doi:10.3109/0142159X.2013.804977
Salkind NJ, ed. Encyclopedia of Research Design . Sage Publishing.
Shaughnessy JJ, Zechmeister EB, Zechmeister JS. Research Methods in Psychology . McGraw Hill Education.
By Anabelle Bernard Fournier Anabelle Bernard Fournier is a researcher of sexual and reproductive health at the University of Victoria as well as a freelance writer on various health topics.
- How it works
"Christmas Offer"
Terms & conditions.
As the Christmas season is upon us, we find ourselves reflecting on the past year and those who we have helped to shape their future. It’s been quite a year for us all! The end of the year brings no greater joy than the opportunity to express to you Christmas greetings and good wishes.
At this special time of year, Research Prospect brings joyful discount of 10% on all its services. May your Christmas and New Year be filled with joy.
We are looking back with appreciation for your loyalty and looking forward to moving into the New Year together.
"Claim this offer"
In unfamiliar and hard times, we have stuck by you. This Christmas, Research Prospect brings you all the joy with exciting discount of 10% on all its services.
Offer valid till 5-1-2024
We love being your partner in success. We know you have been working hard lately, take a break this holiday season to spend time with your loved ones while we make sure you succeed in your academics
Discount code: RP23720

Published by Robert Bruce at August 29th, 2023 , Revised On September 5, 2023
Biology Research Topics
Are you in need of captivating and achievable research topics within the field of biology? Your quest for the best biology topics ends right here as this article furnishes you with 100 distinctive and original concepts for biology research, laying the groundwork for your research endeavor.
Table of Contents
Our proficient researchers have thoughtfully curated these biology research themes, considering the substantial body of literature accessible and the prevailing gaps in research.
Should none of these topics elicit enthusiasm, our specialists are equally capable of proposing tailor-made research ideas in biology, finely tuned to cater to your requirements.
Thus, without further delay, we present our compilation of biology research topics crafted to accommodate students and researchers.
Research Topics in Marine Biology
- Impact of climate change on coral reef ecosystems.
- Biodiversity and adaptation of deep-sea organisms.
- Effects of pollution on marine life and ecosystems.
- Role of marine protected areas in conserving biodiversity.
- Microplastics in marine environments: sources, impacts, and mitigation.
Biological Anthropology Research Topics
- Evolutionary implications of early human migration patterns.
- Genetic and environmental factors influencing human height variation.
- Cultural evolution and its impact on human societies.
- Paleoanthropological insights into human dietary adaptations.
- Genetic diversity and population history of indigenous communities.
Biological Psychology Research Topics
- Neurobiological basis of addiction and its treatment.
- Impact of stress on brain structure and function.
- Genetic and environmental influences on mental health disorders.
- Neural mechanisms underlying emotions and emotional regulation.
- Role of the gut-brain axis in psychological well-being.
Cancer Biology Research Topics
- Targeted therapies in precision cancer medicine.
- Tumor microenvironment and its influence on cancer progression.
- Epigenetic modifications in cancer development and therapy.
- Immune checkpoint inhibitors and their role in cancer immunotherapy.
- Early detection and diagnosis strategies for various types of cancer.
Also read: Cancer research topics
Cell Biology Research Topics
- Mechanisms of autophagy and its implications in health and disease.
- Intracellular transport and organelle dynamics in cell function.
- Role of cell signaling pathways in cellular response to external stimuli.
- Cell cycle regulation and its relevance to cancer development.
- Cellular mechanisms of apoptosis and programmed cell death.
Developmental Biology Research Topics
- Genetic and molecular basis of limb development in vertebrates.
- Evolution of embryonic development and its impact on morphological diversity.
- Stem cell therapy and regenerative medicine approaches.
- Mechanisms of organogenesis and tissue regeneration in animals.
- Role of non-coding RNAs in developmental processes.
Also read: Education research topics
Human Biology Research Topics
- Genetic factors influencing susceptibility to infectious diseases.
- Human microbiome and its impact on health and disease.
- Genetic basis of rare and common human diseases.
- Genetic and environmental factors contributing to aging.
- Impact of lifestyle and diet on human health and longevity.
Molecular Biology Research Topics
- CRISPR-Cas gene editing technology and its applications.
- Non-coding RNAs as regulators of gene expression.
- Role of epigenetics in gene regulation and disease.
- Mechanisms of DNA repair and genome stability.
- Molecular basis of cellular metabolism and energy production.
Research Topics in Biology for Undergraduates
- 41. Investigating the effects of pollutants on local plant species.
- Microbial diversity and ecosystem functioning in a specific habitat.
- Understanding the genetics of antibiotic resistance in bacteria.
- Impact of urbanization on bird populations and biodiversity.
- Investigating the role of pheromones in insect communication.
Synthetic Biology Research Topics
- Design and construction of synthetic biological circuits.
- Synthetic biology applications in biofuel production.
- Ethical considerations in synthetic biology research and applications.
- Synthetic biology approaches to engineering novel enzymes.
- Creating synthetic organisms with modified functions and capabilities.
Animal Biology Research Topics
- Evolution of mating behaviors in animal species.
- Genetic basis of color variation in butterfly wings.
- Impact of habitat fragmentation on amphibian populations.
- Behavior and communication in social insect colonies.
- Adaptations of marine mammals to aquatic environments.
Also read: Nursing research topics
Best Biology Research Topics
- Unraveling the mysteries of circadian rhythms in organisms.
- Investigating the ecological significance of cryptic coloration.
- Evolution of venomous animals and their prey.
- The role of endosymbiosis in the evolution of eukaryotic cells.
- Exploring the potential of extremophiles in biotechnology.
Biological Psychology Research Paper Topics
- Neurobiological mechanisms underlying memory formation.
- Impact of sleep disorders on cognitive function and mental health.
- Biological basis of personality traits and behavior.
- Neural correlates of emotions and emotional disorders.
- Role of neuroplasticity in brain recovery after injury.
Biological Science Research Topics:
- Role of gut microbiota in immune system development.
- Molecular mechanisms of gene regulation during development.
- Impact of climate change on insect population dynamics.
- Genetic basis of neurodegenerative diseases like Alzheimer’s.
- Evolutionary relationships among vertebrate species based on DNA analysis.
Biology Education Research Topics
- Effectiveness of inquiry-based learning in biology classrooms.
- Assessing the impact of virtual labs on student understanding of biology concepts.
- Gender disparities in science education and strategies for closing the gap.
- Role of outdoor education in enhancing students’ ecological awareness.
- Integrating technology in biology education: challenges and opportunities.
Biology-Related Research Topics
- The intersection of ecology and economics in conservation planning.
- Molecular basis of antibiotic resistance in pathogenic bacteria.
- Implications of genetic modification of crops for food security.
- Evolutionary perspectives on cooperation and altruism in animal behavior.
- Environmental impacts of genetically modified organisms (GMOs).
Biology Research Proposal Topics
- Investigating the role of microRNAs in cancer progression.
- Exploring the effects of pollution on aquatic biodiversity.
- Developing a gene therapy approach for a genetic disorder.
- Assessing the potential of natural compounds as anti-inflammatory agents.
- Studying the molecular basis of cellular senescence and aging.
Biology Research Topic Ideas
- Role of pheromones in insect mate selection and behavior.
- Investigating the molecular basis of neurodevelopmental disorders.
- Impact of climate change on plant-pollinator interactions.
- Genetic diversity and conservation of endangered species.
- Evolutionary patterns in mimicry and camouflage in organisms.
Biology Research Topics for Undergraduates
- Effects of different fertilizers on plant growth and soil health.
- Investigating the biodiversity of a local freshwater ecosystem.
- Evolutionary origins of a specific animal adaptation.
- Genetic diversity and disease susceptibility in human populations.
- Role of specific genes in regulating the immune response.
Cell and Molecular Biology Research Topics
- Molecular mechanisms of DNA replication and repair.
- Role of microRNAs in post-transcriptional gene regulation.
- Investigating the cell cycle and its control mechanisms.
- Molecular basis of mitochondrial diseases and therapies.
- Cellular responses to oxidative stress and their implications in ageing.
These topics cover a broad range of subjects within biology, offering plenty of options for research projects. Remember that you can further refine these topics based on your specific interests and research goals.
Frequently Asked Questions
What are some good research topics in biology?
A good research topic in biology will address a specific problem in any of the several areas of biology, such as marine biology, molecular biology, cellular biology, animal biology, or cancer biology.
A topic that enables you to investigate a problem in any area of biology will help you make a meaningful contribution.
How to choose a research topic in biology?
Choosing a research topic in biology is simple.
Follow the steps:
- Generate potential topics.
- Consider your areas of knowledge and personal passions.
- Conduct a thorough review of existing literature.
- Evaluate the practicality and viability.
- Narrow down and refine your research query.
- Remain receptive to new ideas and suggestions.
Who Are We?
For several years, Research Prospect has been offering students around the globe complimentary research topic suggestions. We aim to assist students in choosing a research topic that is both suitable and feasible for their project, leading to the attainment of their desired grades. Explore how our services, including research proposal writing , dissertation outline creation, and comprehensive thesis writing , can contribute to your college’s success.
You May Also Like
Learn how to write an exceptional research paper with the help of professionally crafted research paper examples. Become an expert now!
Explore the essential elements in choosing effective control variables for robust and valid research outcomes.
Craft a compelling scholarship motivation letter by showcasing your passion, achievements, and future goals concisely and impactfully.
Ready to place an order?
USEFUL LINKS
Learning resources.

COMPANY DETAILS

- How It Works
- Methodology
- Research Methodology
- Qualitative Research
What Is Qualitative Research? An Overview and Guidelines
- Australasian Marketing Journal (AMJ)
- This person is not on ResearchGate, or hasn't claimed this research yet.
Discover the world's research
- 25+ million members
- 160+ million publication pages
- 2.3+ billion citations
No full-text available
To read the full-text of this research, you can request a copy directly from the author.

- Mina Seraj-Aksit

- J Consum Behav

- Margaret M. Cullen

- Rossella Gambetti

- Luisella Bovera

- Qual Res Psychol

- Weng Marc Lim

- Karyn Cooper
- Gaurav Gupta

- Bronislaw Malinowski
- QUAL HEALTH RES
- Wendy Duggleby

- Edward F. Fern
- J BUS IND MARK

- Kathy Charmaz
- INT J MANAG REV
- Malvina Klag
- Ann Langley
- Roy Suddaby
- INT J INTERCULT REL
- Yvonna S. Lincoln
- Anselm L. Strauss
- Int J Retail Distrib Manag

- J SOC ISSUES
- Egon G. Guba
- LANDSCAPE URBAN PLAN

- Susan Spiggle
- L Cavusoglu
- Recruit researchers
- Join for free
- Login Email Tip: Most researchers use their institutional email address as their ResearchGate login Password Forgot password? Keep me logged in Log in or Continue with Google Welcome back! Please log in. Email · Hint Tip: Most researchers use their institutional email address as their ResearchGate login Password Forgot password? Keep me logged in Log in or Continue with Google No account? Sign up
- Resources Home
- All Publications
- Digital Resources
- The BERA Podcast
- Research Intelligence
- BERA Journals
- Ethics and guidance
- BERA Blog Home
- About the blog
- Blog special issues
- Blog series
- Submission policy
- Opportunities Home
- Awards and Funding
- Communities Home
- Special Interest Groups
- Early Career Researcher Network
- Events Home
- BERA Conference 2024
- Upcoming Events
- Past Events
- Past Conferences
- BERA Membership
Virtual interviews: A new norm in qualitative research during the Covid-19 pandemic
Olanrewaju Zaid, PhD researcher at University of Bradford Mohammed Abdullahi, Doctoral researcher at University of Warwick 7 Aug 2024
Virtual interviews have gained prominence among researchers from various fields as an effective method for qualitative data collection. The Covid-19 pandemic – with its movement restrictions, isolation measures and the contagious nature of the virus – has driven the transition from traditional face-to-face interviews to virtual or remote interviews using voice-over-internet-protocol (VoIP) technologies such as Microsoft Teams and Zoom.
The rise of remote work during the pandemic has solidified virtual interviews (VIs) as a key method for data collection. VIs closely resemble traditional in-person interviews, involving tasks such as recruiting suitable interviewees and scheduling interviews to avoid conflicts. Relying on VIs also requires a basic level of technological proficiency, including familiarity with computer or mobile phone operation.
The use of technology in virtual interviews is not new in qualitative research and has been considered a viable alternative to in-person interviews (Nehls et al., 2015; Sah et al., 2020). Scholars agree that the ethical considerations for virtual qualitative research are similar to those of in-person research (Roberts et al., 2021). However, VIs introduce an additional layer of security to verify the identities of both the interviewer and interviewee, ensuring the intended individuals participate and that the interview is conducted on a secure network to prevent data breaches.
‘Virtual interviews introduce an additional layer of security to verify the identities of both the interviewer and interviewee, ensuring the intended individuals participate and that the interview is conducted on a secure network to prevent data breaches.’
A set of ten fundamental principles has been proposed to ensure successful virtual interviews for both interviewees and interviewers (Chan et al., 2021). These principles are universally applicable and cover both human and technical aspects, including maintaining professionalism, addressing technology usability, creating a conducive environment, managing environmental factors, adopting a dual-role perspective, meticulous planning, clear communication, active engagement, flexibility, and the use of follow-up emails for gratitude or clarification.
Benefits of virtual interviews
Virtual interviews offer several advantages, highlighted during the Covid-19 pandemic. They provide scheduling flexibility, eliminate the need for physical travel, reduce time constraints, and mitigate health risks. One key benefit is the natural recording and transcription capabilities of certain software packages such as Zoom and Microsoft Teams (Singh et al., 2022). These platforms also provide alerts to remind participants of upcoming sessions. VIs have facilitated participation for individuals with mobility challenges and have streamlined international research involving locations with restricted access (Saarijärvi & Bratt, 2021).
Challenges of virtual interviews for both interviewers and interviewees
Despite their advantages, virtual interviews pose challenges that require investments in high-quality technology, testing and user training (Gray et al., 2020). They sometimes lack the non-verbal cues and body language essential for capturing lived experiences. Bias and engagement are significant challenges, as interviewees often participate from their comfortable environments, which may display personal artefacts or symbols such as political or religious paraphernalia. This comfort zone can lead to distractions, affecting response quality. Additionally, the auto-transcription feature offered by VIs may not always be accurate, and interviewers may become less familiar with the data (Sah et al., 2020).
Exploring the future of virtual interviews
A promising avenue for research is investigating how factors such as artificial intelligence, ergonomics, spatial disparities and virtual augmented reality may impact VoIP in virtual interview technologies across diverse geographical locations.
Chan, D., Fitzsimmons, C. M., Mandler, M. D., & Batista, P. J. (2021). Ten simple rules for acing virtual interviews. PLOS Computational Biology , 17 (6), e1009057. https://doi.org/10.1371/journal.pcbi.1009057
Gray, L. M., Wong-Wylie, G., Rempel, G. R., & Cook, K. (2020). Expanding qualitative research interviewing strategies: Zoom video communications. The Qualitative Report , 25 (5), 1292–1301. https://doi.org/10.46743/2160-3715/2020.4212
Nehls, K., Smith, B. D., & Schneider, H. (2015). Video-conferencing interviews in qualitative research. In S. Hai-Jew (Ed.), Enhancing qualitative and mixed methods research with technology (pp. 140–157). IGI Global.
Roberts, J. K., Pavlakis, A. E., & Richards, M. P. (2021). It’s more complicated than it seems: Virtual qualitative research in the COVID-19 era. International Journal of Qualitative Methods , 20 , https://doi.org/10.1177/16094069211002959
Saarijärvi, M., & Bratt, E. L. (2021). When face-to-face interviews are not possible: Tips and tricks for video, telephone, online chat, and email interviews in qualitative research. European Journal of Cardiovascular Nursing , 20 (4), 392–396. https://doi.org/10.1093/eurjcn/zvab038
Sah, L. K., Singh, D. R., & Sah, R. K. (2020). Conducting qualitative interviews using virtual communication tools amid COVID-19 pandemic: A learning opportunity for future research. Journal of Nepal Medical Association, 58 (232), 1103–1106. https://doi.org/10.31729/jnma.5738
Singh, H., Tang, T., Thombs, R., Armas, A., Nie, J. X., Nelson, M. L., & Gray, C. S. (2022). Methodological insights from a virtual, team-based rapid qualitative method applied to a study of providers’ perspectives of the COVID-19 pandemic impact on hospital-to-home transitions. International Journal of Qualitative Methods , 21 , 16094069221107144. https://doi.org/10.1177/16094069221107144
More related content
Reports 6 Aug 2024
Publication series
News 2 Aug 2024
News 1 Jul 2024
News 25 Jun 2024
News 21 Jun 2024
Thank you for visiting nature.com. You are using a browser version with limited support for CSS. To obtain the best experience, we recommend you use a more up to date browser (or turn off compatibility mode in Internet Explorer). In the meantime, to ensure continued support, we are displaying the site without styles and JavaScript.
- View all journals
- Explore content
- About the journal
- Publish with us
- Sign up for alerts
- Open access
- Published: 08 August 2024
Spermidine is essential for fasting-mediated autophagy and longevity
- Sebastian J. Hofer ORCID: orcid.org/0000-0002-0756-0014 1 , 2 , 3 , 4 , 5 ,
- Ioanna Daskalaki 6 , 7 ,
- Martina Bergmann 1 ,
- Jasna Friščić 8 , 9 ,
- Andreas Zimmermann ORCID: orcid.org/0000-0003-2249-944X 1 , 2 , 3 ,
- Melanie I. Mueller ORCID: orcid.org/0000-0002-7472-2677 1 ,
- Mahmoud Abdellatif ORCID: orcid.org/0000-0002-5042-9054 3 , 4 , 5 , 10 ,
- Raffaele Nicastro ORCID: orcid.org/0000-0002-5420-2228 11 ,
- Sarah Masser ORCID: orcid.org/0000-0002-5195-0650 3 , 12 ,
- Sylvère Durand 4 , 5 ,
- Alexander Nartey 1 ,
- Mara Waltenstorfer ORCID: orcid.org/0000-0001-7541-8338 1 ,
- Sarah Enzenhofer ORCID: orcid.org/0009-0006-7474-5226 1 ,
- Isabella Faimann 1 ,
- Verena Gschiel ORCID: orcid.org/0000-0002-1090-9010 1 ,
- Thomas Bajaj 13 ,
- Christine Niemeyer 13 ,
- Ilias Gkikas 6 , 7 ,
- Lukas Pein 1 ,
- Giulia Cerrato 4 , 5 ,
- Hui Pan 4 , 5 ,
- YongTian Liang ORCID: orcid.org/0000-0002-5802-1045 14 , 15 ,
- Jelena Tadic ORCID: orcid.org/0000-0001-6232-5378 1 , 2 , 3 ,
- Andrea Jerkovic ORCID: orcid.org/0000-0002-0442-712X 1 ,
- Fanny Aprahamian 4 , 5 ,
- Christine E. Robbins 16 ,
- Nitharsshini Nirmalathasan 4 , 5 ,
- Hansjörg Habisch ORCID: orcid.org/0000-0001-5537-506X 17 ,
- Elisabeth Annerer 12 ,
- Frederik Dethloff 18 ,
- Michael Stumpe 11 ,
- Franziska Grundler 19 ,
- Françoise Wilhelmi de Toledo 19 ,
- Daniel E. Heinz 13 ,
- Daniela A. Koppold 20 , 21 , 22 ,
- Anika Rajput Khokhar ORCID: orcid.org/0000-0002-0135-9643 20 , 23 ,
- Andreas Michalsen 20 , 22 ,
- Norbert J. Tripolt 24 , 25 ,
- Harald Sourij 24 , 25 ,
- Thomas R. Pieber 3 , 24 , 25 , 26 ,
- Rafael de Cabo ORCID: orcid.org/0000-0002-3354-2442 27 ,
- Mark A. McCormick 16 ,
- Christoph Magnes ORCID: orcid.org/0000-0002-5153-7444 26 ,
- Oliver Kepp ORCID: orcid.org/0000-0002-6081-9558 4 , 5 ,
- Joern Dengjel ORCID: orcid.org/0000-0002-9453-4614 11 ,
- Stephan J. Sigrist ORCID: orcid.org/0000-0002-1698-5815 14 , 15 ,
- Nils C. Gassen ORCID: orcid.org/0000-0002-4265-3398 13 ,
- Simon Sedej ORCID: orcid.org/0000-0002-4419-6821 3 , 10 , 28 ,
- Tobias Madl ORCID: orcid.org/0000-0002-9725-5231 3 , 17 ,
- Claudio De Virgilio ORCID: orcid.org/0000-0001-8826-4323 11 ,
- Ulrich Stelzl ORCID: orcid.org/0000-0003-2500-3585 2 , 3 , 12 ,
- Markus H. Hoffmann 8 , 9 ,
- Tobias Eisenberg ORCID: orcid.org/0000-0003-3559-1130 1 , 2 , 3 ,
- Nektarios Tavernarakis ORCID: orcid.org/0000-0002-5253-1466 6 , 29 na1 ,
- Guido Kroemer ORCID: orcid.org/0000-0002-9334-4405 4 , 5 , 30 na1 &
- Frank Madeo ORCID: orcid.org/0000-0002-5070-1329 1 , 2 , 3 na1
Nature Cell Biology ( 2024 ) Cite this article
Metrics details
- Macroautophagy
- Metabolomics
- Molecular biology
Caloric restriction and intermittent fasting prolong the lifespan and healthspan of model organisms and improve human health. The natural polyamine spermidine has been similarly linked to autophagy enhancement, geroprotection and reduced incidence of cardiovascular and neurodegenerative diseases across species borders. Here, we asked whether the cellular and physiological consequences of caloric restriction and fasting depend on polyamine metabolism. We report that spermidine levels increased upon distinct regimens of fasting or caloric restriction in yeast, flies, mice and human volunteers. Genetic or pharmacological blockade of endogenous spermidine synthesis reduced fasting-induced autophagy in yeast, nematodes and human cells. Furthermore, perturbing the polyamine pathway in vivo abrogated the lifespan- and healthspan-extending effects, as well as the cardioprotective and anti-arthritic consequences of fasting. Mechanistically, spermidine mediated these effects via autophagy induction and hypusination of the translation regulator eIF5A. In summary, the polyamine–hypusination axis emerges as a phylogenetically conserved metabolic control hub for fasting-mediated autophagy enhancement and longevity.
Continuous caloric restriction (CR) remains the gold standard for extending the lifespan and healthspan of model organisms 1 , 2 . Recently, intermittent fasting (IF) interventions, often combined with CR, emerged as alternatives for clinical implementation 3 . However, to date, it remains uncertain whether IF offers health benefits due to the temporary cessation of caloric intake (without CR) or due to a net reduction of total calories de facto resulting in CR 4 , 5 , 6 . IF, like CR, delays hallmarks of aging in yeast, worms, insects and mice 3 , 7 , 8 , 9 . In humans, intermittent 7 and long-term 10 fasting, as well as continuous CR 11 , are associated with favourable effects on multiple health-relevant parameters that may share a common mechanistic basis. Strong evidence exists that macroautophagy (hereafter referred to as ‘autophagy’) mediates these effects 12 .
In mammals, an age-associated reduction in autophagic flux 13 contributes to the accumulation of protein aggregates and dysfunctional organelles, failing pathogen elimination and exacerbated inflammation 14 . Genetic autophagy inhibition accelerates aging processes in mice 13 and loss-of-function mutations of genes that regulate or execute autophagy have been causally linked to cardiovascular, infectious, neurodegenerative, metabolic, musculoskeletal, ocular and pulmonary diseases, many of which resemble premature aging 15 , 16 , 17 , 18 . Conversely, genetic autophagy stimulation promotes healthspan and lifespan in model organisms, including flies 19 and mice 20 , 21 . Besides nutritional interventions, administering the natural polyamine spermidine (SPD) to yeast, worms, flies and mice is another strategy to extend the lifespan in an autophagy-dependent fashion 22 , 23 , 24 , 25 , 26 . Moreover, SPD restores autophagic flux in circulating lymphocytes from aged humans 25 , 27 , coinciding with the observation that increased dietary SPD uptake is associated with reduced overall mortality in human populations 28 .
Hence, fasting, CR and SPD extend the lifespan of model organisms and activate phylogenetically conserved, autophagy-dependent geroprotection. Intrigued by these premises, we investigated whether the geroprotective effects of IF might be connected to, or depend on, SPD.
Fasting elevates spermidine levels
To investigate polyamine metabolism during IF, we subjected four different organisms to acute fasting stimuli. Mass spectrometry (MS)-based quantification of SPD, the primary biologically active polyamine and its precursors ornithine (ORN), putrescine (PUT), as well as its metabolite spermine (SPM) (Fig. 1a ) revealed a uniform increase in polyamine content upon fasting across various species.

a , Schematic overview of the polyamine pathway and adjacent metabolites. AdoMetDC, adenosylmethionine decarboxylase; ARG, arginase; MAT, methionine adenosyltransferase; ODC, ornithine decarboxylase; SAT, SPD/SPM acetyltransferase, SRM, spermidine synthase; SMOX, spermine oxidase; SMS, spermine synthase; PAO, polyamine oxidase. b , Polyamine levels of WT BY4741 yeast shifted to nitrogen-deprived medium (−N) for the indicated times. Data are normalized to the mean of the control (CTL) condition at every time point. Note that the statistics were performed together with additional groups as indicated in Supplementary Fig. 1a . n = 6 biologically independent samples (yeast cultures). c , WT BY4741 yeast cells were pre-labelled with 13 C 6 -arginine [CTL(Arg*)] and shifted to CTL(Arg*) or −N medium for 6 h. MS-based analysis of labelled products revealed a uniform increase in percentage of labelled polyamines in N-starved cells. nd, not detected. n = 6 biologically independent samples (yeast cultures). d , Polyamine levels of young female w 1118 flies fasted for 12 or 24 h (starting at 20:00 upon lights turned off). Data are normalized to the ad libitum (ad lib) group at every time point. Re-fed, 12 h re-feeding after 24 h fasting. n = 5 (re-fed, 24 h ad lib ORN), 6 (24 h ad lib PUT, SPD and SPM), 7 (rest) biologically independent samples (groups of flies). e , Serum polyamine levels of young male and female C57BL/6 mice fasted or kept ad lib for 14–16 h overnight, starting at 16:00–17:00). n = 5 (male), 8 (female) mice. f , Relative polyamine levels in human serum from cohort 1 after fasting (9 (7–13) days) depicted as mean with s.e.m. and violin plots, showing median and quartiles as lines. The extra panel depicts the individual increase in SPD levels for every volunteer. n = 104 (PUT), 109 (SPD baseline), 100 (SPM baseline), 105 (SPD fasted), 94 (SPM fasted) volunteers. g , Relative polyamine levels in human serum from cohort 2 after increasing numbers of fasting days. n = 61 (baseline PUT), 62 (baseline ORN and SPD), 22 (4–5 d PUT and SPD), 25 (4–5 d ORN, 6–8 d ORN), 19 (6–8 d PUT), 20 (6–8 d SPD), 9 (9–16 d PUT and SPD), 13 (9–16 d ORN) volunteers. h , Relative polyamine and precursor levels in human serum and PBMCs from cohort 4 after increasing numbers of fasting days. BL, baseline, RF = days 3 or 7 after re-introduction of food. N (serum) = 7 (BL SAM), 11 (3 d SAM), 12 (7d RF SAM), 13 (5d SAM), 14 (BL PUT and SPD), 15 (BL ARG and MET; 5 d ARG; 3d RF SAM), 16 (BL ORN; 5 d MET, ORN, PUT and SPD), 17 (3 d ARG, MET, ORN, PUT and SPD; 7 d RF ARG, MET, ORN, PUT and SPD), 18 (3 d RF ARG, MET, ORN, PUT and SPD) volunteers. N (PBMCs) = 6 (5 d ORN), 7 (5 d rest), 9 (BL SPD), 10 (BL rest), 11 (3 d RF SPD) 12 (3 d SPD and SPM; 3 d RF rest), 13 (3 d rest; 7 d RF) volunteers. Statistics used were two-way analysis of variance (ANOVA) with Holm-Šídák’s multiple comparisons test ( b , d , f , h ) and two-tailed Student’s t -tests ( c ). For every analyte ( e ) two-way ANOVA with false discovery rate (FDR) correction (two-stage step-up method by Benjamini, Krieger and Yekutieli, Q = 0.05) together with data depicted in Extended Data Fig. 1f–i (male) and Extended Data Fig. 1k–n (female). Wilcoxon matched-pairs signed rank test ( f ). Kruskal–Wallis test with Dunn’s multiple comparison test ( g ). FC, fold change to control. Heatmaps show means. Bar and line graphs show mean ± s.e.m. * P < 0.05, ** P < 0.01, *** P < 0.001, # P < 0.2. Source numerical data are available in source data.
Source data
Starving Saccharomyces cerevisiae (yeast) cells in water increased the levels of SPD and SPM (Extended Data Fig. 1a ), similar to glucose restriction (Extended Data Fig. 1b ), while ORN generally decreased. Nitrogen starvation (−N), a classical autophagy-inducing intervention 29 , 30 , elicited a fast and transient increase in polyamines, mainly PUT, accompanied by a drastic decrease in the precursor ORN (Fig. 1b ). To shed light on the dynamics under −N, we studied metabolic flux in yeast using 13 C 6 -labelled arginine (ARG). We found that the cellular levels of ARG-derived polyamines after 6 h of nitrogen deprivation were higher than in N-containing control medium (Fig. 1c ). Thus, despite the elimination of extracellular nitrogen, polyamine flux remained active, favouring the utilization of residual ARG molecules for polyamine synthesis.
SPD increased in female and male Drosophila melanogaster (fruit fly) fasted for 24 h (Fig. 1d and Extended Data Fig. 1c ), which caused a body weight loss of 10% and 5%, respectively (Extended Data Fig. 1d ). This SPD increase was reversed by 12 h re-feeding (Fig. 1d and Extended Data Fig. 1c ). Similarly, young male and female C57BL6/J mice fasted for 14–16 h, which caused significant weight loss (Extended Data Fig. 1e,j ), had higher SPD levels (but not PUT nor SPM) in the serum than their ad libitum-fed controls, whereas ORN content was reduced (Fig. 1e ). Moreover, the abundance of ORN and polyamines changed significantly in several organs of acutely fasted mice in a tissue-specific manner (Extended Data Fig. 1f–i,k–n ), favouring an increase of SPD in multiple tissues. We next asked whether these alterations in polyamine content would also occur during long-term CR, starting at 9 months of age 31 . We found increased serum SPD at 17 months of age in male, but not female, mice (Extended Data Fig. 1o,p ). At later time points (21 months), female CR mice also showed significantly elevated PUT and SPD levels in skeletal muscle (Extended Data Fig. 1q ).
Furthermore, starvation elevated SPD and SPM levels uniformly in human U2OS osteosarcoma and H4 glioblastoma cells (Extended Data Fig. 1r,s ). Accordingly, in nutrient-depleted U2OS cells, the expression of arginase ( ARG1 ), ornithine decarboxylase ( ODC1) , spermidine/spermine N1-acetyltransferase 1 ( SAT1 ) and glycine N -methyltransferase ( GNMT ) increased compared with cells cultured in control medium, whereas the polyamine-associated transcription factors MYC and YAP/TAZ , as well as two recently identified polyamine transporters, ATPase cation transporting 13A2/13A3 ( ATP13A2/3 ), were unaffected (Extended Data Fig. 1t ).
In human volunteers, long-term therapeutic fasting with a daily caloric intake of approximately 250 kcal (Supplementary Table 1 ) under clinical supervision for 7–13 days, SPD levels (but not PUT nor SPM) significantly increased in the serum (Fig. 1f ). This increase in SPD content was similarly found in men and women (Extended Data Fig. 1u ), and was independent of age and body mass index (BMI) before the intervention or body weight loss (Extended Data Fig. 1v–x ).
We analysed an independent cohort of volunteers fasting for variable periods (Supplementary Table 1 ; cohort 2). SPD levels increased by ~50% after 4–5 days and remained elevated during long-term fasting (Fig. 1g ). In a third cohort (Supplementary Table 1 ), we analysed plasma SPD of individuals who voluntarily followed an IF routine (12-h eating periods followed by 36-h zero-calorie periods for several months). Again, we observed an elevation in SPD levels (Extended Data Fig. 1y ). Finally, SPD increased in serum and peripheral blood mononuclear cells (PBMCs) of a separate, fourth cohort of volunteers during fasting and reverted to baseline levels after re-feeding (Fig. 1h ). Notably, in these PBMCs, ODC1 protein levels decreased during fasting (Extended Data Fig. 1z ), suggesting that elevated SPD levels either caused a feedback repression of ODC1 or, at least in these cells, might stem from increased uptake rather than intracellular synthesis.
In conclusion, nutrient starvation (yeast and human cell lines), overnight fasting (flies, mice and humans), long-term CR (mice) or long-term fasting (humans) induced SPD elevation.
Polyamine synthesis is required for efficient metabolic remodelling and TORC1 inhibition during fasting
Next, we investigated the cellular consequences of impaired polyamine anabolism on acute fasting responses. We generated yeast lacking the rate-limiting enzyme ornithine decarboxylase (ODC1; yeast Spe1), which are characterized by polyamine depletion (Supplementary Fig. 1a ). This strain (∆ spe1 ) showed no elevation of polyamines upon starvation, and SPD supplementation (100 µM) fully replenished the intracellular SPD pool (Fig. 2a ). We subjected ∆ spe1 cells with and without SPD to proteomic analyses after 6 h nitrogen starvation (Supplementary Fig. 1b ). Principal-component analysis (PCA) of the proteome revealed a clear distinction between the genotypes and that SPD could revert ∆ spe1 -associated global differences, whereas it did not affect the wild-type (WT) proteome (Fig. 2b ). Mapping the identified proteins to Kyoto Encyclopedia of Genes and Genomes (KEGG) terms, we found several pathways that have been implicated in the starvation response dysregulated in ∆ spe1 (Supplementary Fig. 1c ). This included, for example, the metabolism of several amino acids (including ARG), lipids and fatty acids, as well as energy-relevant pathways (tricarboxylic acid (TCA) cycle and oxidative phosphorylation). Notably, we also found a disturbed starvation response of the proteostasis-associated pathways autophagy and TORC1/2, the yeast homologues of mechanistic target of rapamycin complex 1/2 (mTORC1/2), in ∆ spe1 versus WT cells (Fig. 2c and Supplementary Fig. 1c ).

a , Relative SPD levels of WT and ∆ spe1 yeast cells after SPD treatment (100 µM) and N starvation. n = 6 biologically independent samples (yeast cultures). b , PCA depicting the proteome change in WT and Δ spe 1 cells under specified condition treatments following a 6-h culture in control or −N medium with or without 100 µM SPD. PCA was performed on a singular-value decomposition of centred and scaled protein groups ( n = 4,684) displays a comparison between PC1 and PC2, along with the representation of a 95% confidence interval for each group. n = 6 biologically independent samples (yeast cultures). c , Differential expression ( z -score) of proteins involved in the TORC complex, from the proteome analysis shown in b . n = 6 biologically independent samples (yeast cultures). d , Volcano plot showing significantly different intracellular metabolites in WT or ∆ spe1 after 6 h −N compared with control conditions. Venn diagram showing exclusive and overlapping significantly regulated metabolites. FDR-corrected P value < 0.05, FC > 1.5. n = 4 biologically independent samples (yeast cultures). e , Metabolomic disturbances in ∆ spe1 cells are rescued by SPD (100 µM) supplementation. The PCA displays a comparison between PC1 and PC2, along with the representation of a 95% confidence interval for each group. n = 6 biologically independent samples (yeast cultures). f , SPD (100 µM) corrects ∆ spe1 -associated amino acid disturbances. Relative arginine and serine levels from metabolomics analysis shown in e . n = 6 biologically independent samples (yeast cultures). Statistics were two-way ANOVA with Holm-Šídák’s multiple comparisons test ( a , f ) and two-tailed Student’s t -tests with FDR correction ( d ). Bar graphs show the mean ± s.e.m. Source numerical data are available in source data.
The prominent dysregulation in metabolic pathways was supported by unbiased metabolomic profiling by nuclear magnetic resonance (NMR) spectroscopy, revealing substantial differences in the intracellular metabolomes after nitrogen deprivation (Supplementary Fig. 2 ). The metabolic disturbances affecting starved ∆ spe1 cells (Fig. 2d ) confirmed findings from the proteome analysis, including increased citric acid and reduced levels of nicotinamide adenine dinucleotide (NAD + ) and adenosine/guanosine-X-phosphate (AXP/GXP; where X stands for mono-, di- or triphosphate), suggesting a disrupted energy metabolism secondary to the loss of intracellular polyamine synthesis (Supplementary Fig. 2 ). Our analysis also indicated dysregulated amino acid homoeostasis, which is normally maintained by autophagy in starving yeast 32 , 33 , 34 .
Notably, exogenously supplemented SPD reversed the metabolic dysregulations in ∆ spe1 cells, both in control and nitrogen-starvation medium, whereas it hardly affected the general WT metabolomes (Fig. 2e and Extended Data Fig. 2a,b ). This included normalized amino acid metabolism (Fig. 2f and Extended Data Fig. 2c,d ), which has been critically linked to TOR and autophagy regulation (for example, for arginine 35 and serine 33 ) and metabolites central to energy metabolism (glucose, NAD + and citric acid, among others) (Fig. 2f and Extended Data Fig. 2e ). Overall, Spe1 was required for the metabolic switch from glycolysis to oxidative phosphorylation, a key event in the cellular adaptation to nitrogen starvation, which partly depends on functional autophagy 36 . For instance, glucose levels were less increased under −N, and the ratios of TCA cycle metabolites were heavily dysregulated (for example, citric acid to succinic acid) (Extended Data Fig. 2e ).
Given the implication of TORC1 in the fasting response of ∆ spe1 yeast cells, we next asked whether ∆ spe1 cells would functionally alter the nitrogen deprivation-induced inhibition of TORC1. Sch9 (the yeast equivalent of mammalian p70 S6K ) dephosphorylation, was significantly delayed in ∆spe1 cells (Fig. 3a,b ). Focusing on TORC-associated proteins in our proteome data, we found dysregulated TORC subunits, including Tco89, Avo1/2 and Tsc11, under both conditions in the ∆ spe1 strain (Extended Data Fig. 3a ). Of note, low levels of polyamines (100 µM) completely reverted the delayed TORC1 inhibition in ∆spe1 cells (Fig. 3c,d ). On the other hand, high levels of additional SPD (5 mM) did not affect TORC1 activity in the WT strain (Fig. 3e,f ). Similar to −N, acute pharmacological inhibition of TORC1 with rapamycin led to a rapid increase of ORN (likely due to the known activation of arginase expression 37 , 38 ) as well as elevated SPD and SPM levels (Extended Data Fig. 3b ). Thus, the efficient shutdown of yeast TOR signalling upon −N, a key event for autophagy induction 39 , requires intact polyamine metabolism, whereas TOR inhibition promotes the anabolism of polyamines.

a , Decrease of TORC1 activity as inferred by Sch9-phosphorylation during -N in WT and ∆ spe1 cells. Representative immunoblot. b , Quantification of immunoblots as shown in a . n = 3 biologically independent samples (yeast cultures). c , Polyamine supplementation (100 µM) corrects the delayed decrease of TORC1 activity during −N in ∆ spe1 cells. Representative immunoblot. d , Quantification of immunoblots as shown in c . n = 4 biologically independent samples (yeast cultures). e , Supplementation of high SPD levels (5 mM) corrects the delayed decrease of TORC1 activity in ∆ spe1 cells but does not affect TORC1 activity in WT cells. Representative immunoblot. f , Quantification of immunoblots as shown in e . n = 4 biologically independent samples (yeast cultures). g , SPD supplementation (100 µM) corrects decreased Atg7 protein levels ∆ spe1 cells in both control and −N medium, as detected in proteome analysis shown in Fig. 2b . n = 6 biologically independent samples (yeast cultures). h , Representative immunoblots of yeast WT and ∆s pe1 GFP-Atg8 cells after 6 h −N with and without 100 µM SPD, assessed for GFP and GAPDH. i , Quantifications of h . n = 6 biologically independent samples (yeast cultures). j , ALP activity (RFU per µg) from Pho8∆N60 assay normalized to each CTL group after 6 h −N. n = 10 (WT CTL, ∆ spe1 CTL), 11 (WT -N), 8 (∆ spe1 −N) biologically independent samples (yeast cultures). k , Representative immunoblots of yeast WT and ∆s pe1 GFP-Atg8 after 6 h −N, with or without ascending concentrations of SPD, assessed for GFP and GAPDH. l , Quantifications of k . n = 6 biologically independent samples (yeast cultures). m , Representative images of human U2OS GFP-LC3 cells starved for 6 h in Hanks’ balanced salt solution (HBSS) (with or without chloroquine (CQ) for 3 h before fixation) after 3 days of 100 µM DFMO treatment. For quantifications see also Extended Data Fig. 5c . Scale bar, 10 µm. n , Quantification of cytosolic GFP-LC3 dots from l , normalized to the average number of GFP-LC3 dots in the control condition. n = 6 biologically independent experiments. o , Representative images of the head region of young control and odc-1(RNAi) C. elegans MAH215 (sqIs11 [lgg-1p::mCherry::GFP::lgg-1 + rol-6]) (LGG-1 is the C. elegans orthologue of LC3) fasted for two days. Autolysosomes (ALs) appear as mCherry-positive puncta. Autophagic activity is indicated by a shift to the red spectrum due to fluorescence quenching of the pH-sensitive-GFP by the acidic environment of the autolysosome. Scale bar, 50 μm. p , Quantification of ALs as depicted in o . Note that the statistics were performed together with additional groups as indicated in Extended Data Fig. 10c . n = 11 (CTL ad lib), 26 (CTL fasted), 8 ( Odc-1(RNAi) ad lib), 30 ( Odc-1(RNAi) fasted) worms. Statistics used were two-way ANOVA with Holm-Šídák’s multiple comparisons test ( b , d , f , g , h , j , n ), one-way ANOVA with Holm-Šídák’s multiple comparisons test ( l ) and Kruskal–Wallis test with FDR correction (two-stage step-up method by Benjamini, Krieger and Yekutieli, Q = 0.05) ( p ). Bar graphs show the mean ± s.e.m. Source numerical data and unprocessed blots are available in source data. NS, not significant.
To translate these findings in vivo, we measured polyamines and precursors in hearts from young male mice overexpressing the human insulin-like growth factor 1 receptor (IGF1R tg ) 40 or carrying a dominant negative phosphoinositide 3-kinase mutant (dnPI3K) 41 specifically in cardiomyocytes. IGF1R tg causes increased IGF1R signalling, leading to elevated mTOR activity and autophagy inhibition, as well as age-associated heart failure, which can be overridden by SPD 42 . Conversely, the cardiomyocyte-specific dnPI3K mutation inhibits mTOR and enhances autophagic flux 42 . We observed a trend towards lower cardiac SPD levels in IGF1R tg mice ( P = 0.222 ) and significantly elevated levels of SPD in dnPI3K mice (Extended Data Fig. 3c ).
Collectively, blocking polyamine synthesis caused a defective cellular response to −N in yeast, thus compromising, inter alia, energy metabolism and amino acid homoeostasis, both of which interface with TOR signalling, an integral hub for sensing and relaying nutrient information to cellular responses and functional autophagy. SPD supplementation reversed the metabolic inflexibility of ∆s pe1 cells.
Spermidine is vital for autophagy induction during fasting
mTOR is a major repressor of autophagic flux and proteomics revealed a profound dysregulation of autophagy-relevant proteins upon SPE1 loss (Extended Data Fig. 3d–f ). SPD has been previously shown to induce ATG7 expression in yeast 23 , and accordingly, SPE1 knockout caused reduced Atg7 protein expression (Fig. 3g ). Furthermore, several autophagy-relevant proteins were differently modulated in the ∆ spe1 strain upon −N (Extended Data Fig. 3d–f ). This included the transcription factors Gcn4 (for amino acid biosynthesis), Msn4 (stress response), several autophagy-related proteins (Atg2/8/11/16/38), as well as vacuolar proteinases (Prb1, Ysp3 and Pep4) and proteins involved in intracellular vesicle trafficking (Vps33, Arc19, Vti1 and Ykt6), among others.
As a functional consequence, ∆ spe1 cells exhibited reduced autophagy induction in response to −N. We observed a diminished autophagy-dependent proteolytic liberation of green fluorescent protein (GFP) from GFP fused to autophagy-related protein 8 (GFP-Atg8, Atg8 being the yeast orthologue of the mammalian LC3 family) (Extended Data Fig. 4a–d ), which was rescued by SPD supplementation (Fig. 3h,i ). This was confirmed with additional autophagy assays, including the reduced redistribution of GFP-Atg8 towards autophagic vacuoles, compared with WT cells (Extended Data Fig. 4e–g ) and the Pho8∆N60 assay 43 (Fig. 3j and Extended Data Fig. 4h ). However, supplementing SPD could not further elevate autophagic flux under −N in WT cells (Fig. 3k,l ). Similarly, rapamycin-induced autophagy was significantly curtailed in ∆ spe1 cells (Extended Data Fig. 4i–l ), which again could be partly rescued by SPD (Extended Data Fig. 4m,n ).
The ODC1 inhibitor difluoromethylornithine (DFMO) depleted polyamines in human U2OS and H4 in nutrient-rich (Extended Data Fig. 5a ) and starved (Extended Data Fig. 5b ) conditions to <5% of control levels, hence excluding the possibility that ODC1 -independent mechanisms (such as uptake of exogenous SPD) would account for the starvation-induced elevation of SPD. DFMO reduced the translocation of GFP–LC3 to autophagosomes and autolysosomes in response to starvation in the presence of the lysosomal inhibitor chloroquine (CQ), indicating that polyamine synthesis is required for autophagic flux (Fig. 3m,n and Extended Data Fig. 5c–f ). These phenotypes could be rescued by co-treatment with 10 µM SPD (Extended Data Fig. 5g–m ). Three siRNAs targeting ODC1 phenocopied the effects of DFMO, hence depleting intracellular polyamines from U2OS cells (Extended Data Fig. 5n ) and dampening starvation-induced autophagic flux similar to ATG5 siRNA (Extended Data Fig. 5o–q ). As found in yeast, SPD supplementation failed to further enhance autophagy in starved U2OS GFP–LC3 cells (Extended Data Fig. 5r,s ). Moreover, mTORC1 inhibition by rapamycin or torin-1 induced DFMO-inhibitable autophagy in U2OS and H4 cells (Supplementary Fig. 3a–f ), which was rescued by SPD (Supplementary Fig. 3g–l ).
C. elegans subjected to acute fasting exhibited elevated messenger RNA levels of odc-1 , amx-3 and hpo-15 (PAOX and SMOX orthologues), whereas spermidine synthase ( spds-1 ) and the AMD1 orthologue smd-1 decreased. Among the tested polyamine-relevant genes, argn-1 (ARG1/2 orthologue) and d2023.4 (SAT1/2) remained unaffected (Extended Data Fig. 6a ). Knockdown of odc-1 by RNA interference (Extended Data Fig. 6b ) reduced fasting-induced autophagic flux assessed by a tandem-tagged (mCherry/GFP) LGG-1 fluorescent reporter, the worm equivalent of yeast Atg8 and human LC3 (Fig. 3o,p and Extended Data Fig. 6c,d ). This was confirmed in a strain expressing an alternative autophagy-sensitive biosensor, GFP-fused SQST-1 (the worm orthologue of human sequestosome 1 (SQSTM1)/p62, an autophagy substrate), which was less degraded in fasted odc-1 knockdown worms than in control nematodes (Extended Data Fig. 6e,f ). Notably, SPD feeding reverted the autophagic deficit of odc-1 worms (Extended Data Fig. 6g,h ). Similar to yeast, odc-1 knockdown also caused a trend ( P = 0.140) towards reduced number of rapamycin-induced autolysosomes (Extended Data Fig. 6i,j ).
Altogether, these findings indicate that ODC1 is required for optimal fasting-induced autophagy across species.
Spermidine is required for fasting-mediated lifespan extension
Nitrogen deprivation reduces the fraction of dead cells in chronological aging experiments performed on yeast, a model for post-mitotic aging 44 , in an autophagy-dependent manner 45 . This longevity-extending effect was abolished in ∆ spe1 cells, indicating that polyamine synthesis is required for longevity upon −N (Fig. 4a,b ). The knockout of spermidine synthase (∆ spe3 ) and that of SAM decarboxylase (∆ spe2 ), which are both required for SPD generation, phenocopied ∆ spe1 with respect to the loss of the longevity during −N (Fig. 4c ). In contrast, the knockout of spermine synthase (∆ spe4 ) failed to affect survival under nitrogen-deprived conditions (Fig. 4c ). Of note, survival deficits triggered by the loss of Spe1 could be fully rescued by the addition of PUT, SPD or SPM ( P > 0.05 against each other under nitrogen starvation) (Supplementary Fig. 4b ). However, increasing concentrations of SPD only improved the nitrogen starvation-prolonged chronological lifespan of WT cells on early time points (Supplementary Fig. 4c ).

a , Representative microscopy images on day 5 of chronological lifespan experiments of yeast WT and ∆ spe1 in control and −N medium, stained with propidium iodide (PI). Scale bar, 5 µm. b , PI-negative (live) cells during chronological aging of yeast WT and ∆ spe1 in control and −N medium. n = 36 (WT), 26 (∆ spe1 CTL), 24 (∆ spe1 −N) biologically independent samples (yeast cultures). c , PI-negative (live) cells during chronological aging yeast WT and ∆ spe2 , ∆ spe3 and ∆ spe4 cells grown to the log phase and shifted to CTL or −N. n = 8 biologically independent samples (yeast cultures). d , Lifespan of female w 1118 flies fed standard food with or without 10 mM DFMO and subjected to IF 12:12 . n = 315 (ad lib), 313 (IF), 327 (ad lib + DFMO), 348 (IF + DFMO) flies. e , Flies from d were assessed for their climbing ability, measured as covered walking distance after a negative geotaxis stimulus, between days 53–60. n = 11 biologically independent samples. f , Flies from d were assessed for their climbing ability, measured as speed after a negative geotaxis stimulus, between days 53–60. n = 11 biologically independent samples. g , Lifespan of C. elegans N2 fed control (CTL) or odc-1(RNAi) expressing bacteria during IF 48:48 . The worms were transferred every other day. IF groups were transferred to agar plates without bacteria every second day. Note that the statistics and experiments were performed together with the groups depicted in i and Fig. 6p . n = 913 (CTL ad lib), 750 (CTL IF), 794 ( odc-1(RNAi) ad lib), 779 ( odc-1(RNAi) IF) worms. h , The susceptibility of worms to heat stress is reduced by IF 48:48 in control, but not in worms fed odc-1(RNAi) expressing bacteria. Young worms (after the first round of fasting) were placed at 37 °C for 6 h. Survival was assessed after overnight recovery at 20 °C. n = 5 biologically independent experiments. i , Lifespan of C. elegans N2 fed bacteria expression control (CTL) or RNAi against argn-1 , spds-1 or smd-1 during IF 48:48 . Note that the experiments and statistics were performed together with the groups depicted in g and Fig. 6p . n = 913 (CTL ad lib), 750 (CTL IF), 899 ( argn-1(RNAi) ad lib), 797 ( argn-1(RNAi) IF), 820 ( smd-1(RNAi) ad lib), 803 ( smd-1(RNAi) IF), 881 ( spds-1(RNAi) ad lib), 746 ( spds-1(RNAi) IF) worms. Statistics used were two-way ANOVA with Holm-Šídák’s multiple comparisons test ( b , c , e , f , h ) and log-rank test with Bonferroni correction ( d , g , i ). Bar graphs show the mean ± s.e.m. Source numerical data are available in source data. AUC, area under the curve.
We next investigated the involvement of SPE1 in other lifespan-increasing interventions in yeast. Inhibition of TOR with rapamycin extends yeast lifespan in an autophagy-dependent fashion 46 , 47 , and this effect was diminished in the ∆ spe1 strain (Supplementary Fig. 4d ) but rescued by polyamine supplementation (Supplementary Fig. 4e ). The extension of chronological lifespan by glucose restriction was partially compromised by the spe1 knockout (Supplementary Fig. 4f ). Replicative aging, which reflects the diminished replicative capacity of aging mother cells 44 , is especially responsive to glucose restriction. Knockout of SPE1 diminished the survival, median (WT 23 versus ∆s pe1 19 days) and maximal replicative lifespan (68 versus 44 days) in low-glucose (0.05%) cultures but did not affect replicative lifespan when glucose concentrations were kept at standard levels (2%) (Supplementary Fig. 4g ). In summary, SPE1 and SPD are essential for longevity induction by nitrogen starvation, rapamycin and glucose depletion.
Testing these findings’ relevance, we subjected fruit flies to an IF regime that improves healthspan and lifespan. We followed the survival of female and male w 1118 flies under IF 12:12 , during which increased daytime food intake compensated for the nightly calorie loss after 10 cycles and at later time points (Extended Data Fig. 7a ). DFMO lowered whole-body SPD levels (Extended Data Fig. 7b ) and reduced the effects of IF 12:12 on improved survival in both sexes (Fig. 4d , Extended Data Fig. 7c , Supplementary Table 2 ). Generally, IF 12:12 seemed more effective in female flies, which we further tested for age-sensitive locomotor capacity with a modified negative geotaxis assay. DFMO prevented locomotion improvement by IF 12:12 (Fig. 4e,f ). Of note, DFMO did not affect food consumption during the first cycles of IF 12:12 (Extended Data Fig. 7d ) or body weight during acute fasting (Extended Data Fig. 7e ) and generally seemed to be non-toxic for flies. Female flies lacking one functional copy of Odc1 ( Odc1 MI10996 mutant) were also unresponsive to IF 12:12 , and nightly SPD feeding could re-instate IF 12:12 -mediated longevity in such flies (Extended Data Fig. 7f and Supplementary Table 2 ).
In C. elegans , genetic inhibition of odc-1 reduced the lifespan extension (Fig. 4g and Supplementary Table 2 ) and heat stress resistance (Fig. 4h ) conferred by IF 48:48 , but did not affect the body size of fasted worms (Extended Data Fig. 7g ). Knockdown of spds-1 (spermidine synthase) or smd-1 (adenosylmethionine decarboxylase 1), which are critical for SPD synthesis, as well as argn-1 (Extended Data Fig. 7h ) reduced the lifespan extension elicited by IF 48:48 in worms (Fig. 4i and Supplementary Table 2 ). Notably, SPD significantly extended the lifespan of intermittently fasted odc-1 knockdown worms towards that of WT controls (Extended Data Fig. 7i and Supplementary Table 2 ). However, SPD did not modulate the IF 48:48 effect on the lifespan of WT worms and was less potent than IF alone when fed ad libitum (Extended Data Fig. 7i and Supplementary Table 2 ). Similarly, rapamycin-triggered longevity was blunted upon odc-1 knockdown in worms (Extended Data Fig. 7j and Supplementary Table 2 ).
Altogether, these findings unravel a pathway in which endogenous polyamine biosynthesis mediates the lifespan-extending effects of nitrogen starvation in yeast and IF in flies and worms.
Cardioprotective and antiarthritic effects of intermittent fasting are blunted by DFMO feeding in mice
We next determined whether endogenous SPD biosynthesis in mice was essential for IF-induced healthspan improvements, focusing on cardioprotection and suppression of inflammation, knowing that both are elicited by SPD supplementation 24 , 48 .
In aged male mice, DFMO abolished favourable cardiac effects of an IF 16:8 protocol (daily 16 h fasting and 8 h ad libitum food access during the light phase) (Fig. 5a and Extended Data Fig. 8a,b ). This concerned improvements in cardinal signs of cardiac aging 49 , including left ventricular (LV) diastolic dysfunction (E/e′) (Fig. 5b and Supplementary Table 3 ; P = 0.042, ad libitum versus IF), LV hypertrophy (Fig. 5c ; P = 0.043) and LV remodelling index ( P = 0.072) (Extended Data Fig. 8c and Supplementary Table 3 ), whereas other cardiac parameters remained unaffected by DFMO and IF 16:8 (Extended Data Fig. 8d,e and Supplementary Table 3 ). DFMO did not alter the IF 16:8 -induced reduction of body weight and food intake (Extended Data Fig. 8f–h ). In a second cohort of aged male mice, we tested the effects of DFMO on general and muscular healthspan improvements elicited by an IF + CR (30%) combination (IF CR ; one meal a day, provided shortly before the dark phase) (Fig. 5d ) 50 . DFMO prevented IF CR -mediated improvements in a visual frailty index 51 (Fig. 5e ), grip strength (Fig. 5f and Extended Data Fig. 8i ) and wire hanging ability (Extended Data Fig. 8j ), whereas it did not affect body weight (Extended Data Fig. 8k ), body composition (Extended Data Fig. 8l ) or body surface temperature (Extended Data Fig. 8m ). Additionally, in young mice, both IF 24:24 and oral SPD supplementation (Fig. 5g ) ameliorated the progression of autoantibody-induced arthritis, and the antiarthritic effects of IF 24:24 were blunted by parallel DFMO administration (Fig. 5h,i ), with comparable results in female and male mice (Extended Data Fig. 8n ).

a , Experimental layout to study the effects of DFMO feeding on IF 16:8 -mediated cardioprotection in 17–18-month-old male C57BL/6J mice. Mice were fasted from 15:00 to 7:00 (16 h daily, excluding weekends). A subset of aged mice received DFMO in the drinking water. After 10 weeks, cardiac function and structure were assessed by echocardiography. Data are shown in b , c , Extended Data Fig. 8a–h and Supplementary Table 3 . b , Ratio of peak early Doppler transmitral flow velocity (E) to myocardial tissue Doppler velocity (e′), a measure of diastolic dysfunction, in aged male mice treated with IF 16:8 , with or without DFMO ( n = 13, 15, 9 and 8 mice per group, respectively). c , LV mass normalized to body surface area (LV mass i ). n = 13 (ad lib), 15 (IF), 9 (ad lib + DFMO), 8 (IF + DFMO) mice. d , Experimental layout to study the effects of DFMO feeding on IF CR -mediated healthspan improvements in 20-month-old male C57BL/6J mice. Mice were given a single meal per day (30% CR) shortly before the dark phase. A subset of mice received 0.25% or 0.5% DFMO in the drinking water. After 3 months, healthspan was assessed. Data are shown in e , f and Extended Data Fig. 8i–m . e , Visual frailty index of aged male mice treated with IF CR , with or without DFMO. n = 12 (ad lib 0.5%), 13 (IF 0.25%), 14 (ad lib 0.25%), 15 (IF 0%), 16 (IF 0.5%), 18 (ad lib 0%) mice. f , Grip strength of fore limbs in gram-force (gf) normalized to body weight of aged male mice treated with IF CR , with or without DFMO. n = 12 (ad lib 0.5%), 13 (IF 0.25%), 14 (ad lib 0.25%), 15 (IF 0%), 16 (IF 0.5%), 18 (ad lib 0%) mice. g , Experimental layout to study the effects of DFMO feeding on IF-mediated anti-inflammatory effects in young BALB/cJRj mice of both sexes. DFMO and 3 mM SPD were supplemented via drinking water. Mice were fasted every other day. IF 24:24 and SPD treatments were started 3 weeks before serum transfer and continued until the end of the experiment. Data are shown in h , i and Extended Data Fig. 8n . h , Development of arthritis upon injection of serum from K/BxN mice in young male and female BALB/cJRj mice treated as outlined in g . n = 8 mice. i , AUC analysis of h . n = 8 mice. Statistics used were two-way ANOVA with Holm-Šídák’s multiple comparisons test ( b , c , e , f ), log-rank test with Bonferroni correction ( c , e ) and one-way ANOVA with Holm-Šídák’s multiple comparisons test ( i ). Bar graphs show the mean ± s.e.m. M, months of age. Source numerical data are available in source data.
Collectively, genetic or pharmacologic ODC1 inhibition attenuated or abolished the healthspan-promoting, cardioprotective and inflammation-regulating effects of IF regimens in multiple distantly related species. Therefore, SPD is probably a dominant pro-autophagic and anti-aging effector metabolite accounting for the beneficial effects of various forms of IF.
Increased eIF5A hypusination is required for fasting-induced longevity
U2OS GFP–LC3 cells were screened for the effects of small interfering RNAs (siRNAs) targeting genes previously linked to SPD effects that hence might modulate starvation-induced autophagy. Besides the knockdown of ODC1 and the autophagy-essential genes ATG5 and ULK1 , we found that depletion of EIF5A (eukaryotic translation initiation factor 5A) significantly reduced the number of GFP–LC3 dots after starvation (Extended Data Fig. 8o,p ). SPD is required to hypusinate eIF5A via a conserved reaction involving deoxyhypusine synthase (DHS) and deoxyhypusine hydroxylase (DOHH) 52 . The resulting covalently modified and active hypusinated eIF5A (eIF5A H ) is involved in the pro-autophagic and antiaging effects of SPD administration 53 , 54 (Fig. 6a ). We thus investigated whether the polyamine–eIF5A H axis played a role in autophagy and lifespan regulation during −N and IF.

a , Scheme of the hypusination pathway. DHS, deoxyhypusine synthase; DOHH, deoxyhypusine hydroxylase; eIF5A, eukaryotic translation initiation factor 5A-1. b , Representative immunoblot of hypusinated eIF5A (eIF5A H ) levels in yeast GFP-Atg8 WT and ∆ spe1 after 6 h of −N. c , Quantification of b . n = 8(WT), 7(∆ spe1 ) biologically independent samples (yeast cultures). d , Representative maximum projection images of confocal microscopy images of female w 1118 fly central brain regions probed for eIF5A and hypusine by immunofluorescence. Before dissection, the flies were fasted for 0 h (ad lib) or 12 h, starting at 20:00. Scale bar, 50 µm. e , Quantification of signal intensities in d . n = 22 (ad lib), 23 (fasted) fly brains. f , Immunoblots of 48 h fasted C. elegans assessed for hypusine and GAPDH. g , Quantification of f . n = 4 biologically independent samples (worm lysates). h , Representative immunoblot of eIF5A H , eIF5A and GAPDH signals of liver samples from ad lib and fasted (14–16 h) young, male C57BL/6 mice. i , Quantification of h . n = 7 mice. j , eIF5A H and total eIF5A levels in human PBMCs after increasing fasting times, measured by capillary immunoblotting. RF, 3 days after re-introduction of food. n = 17 (days 0, 1 and 3), 15 (day 5), 16 (day 7 and RF) volunteers. k , Representative immunoblot of yeast Lia1-6xHA, assessed for HA-tags and GAPDH after 6 and 24 h −N. l , Quantification of Lia1-6xHA levels as depicted in k . n = 8 biologically independent samples (yeast cultures). m , Quantification of relative mRNA expression of dhps-1 , dohh-1 and iff-1 in 24 h fasted C. elegans . n = 3 ( iff-1 ), 5 ( dhps-1 and dohh-1 ) biologically independent experiments. n , Quantification of relative mRNA expression of DHPS , DOHH and EIF5A in 6 h starved U2OS cells. n = 3 ( DOHH) , 4 ( DHPS and EIF5A ) biologically independent experiments. o , PI-negative (live) yeast cells during chronological lifespan analysis of WT and ∆ lia1 yeast in control and −N medium. n = 12 biologically independent samples (yeast cultures). p , Lifespan of C. elegans N2 fed control (CTL) or dhps-1(RNAi) (homologue of DHS) expressing bacteria during IF 48:48 . Note that the experiments and statistics were performed together with the groups depicted in Fig. 4g,i . n = 913 (CTL ad lib), 750 (CTL IF), 862 ( dhps-1(RNAi) ad lib), 776 ( dhps-1(RNAi) IF) worms. Statistics used were two-way ANOVA with Holm-Šídák’s multiple comparisons test ( c , j , l , o ), two-tailed Student’s t -tests with Holm-Šídák’s multiple comparisons test ( e , i , m , n ), two-tailed Student’s t -test ( h ) and log-rank test with Bonferroni correction ( p ). Bar and line graphs show the mean ± s.e.m. Source numerical data and unprocessed blots are available in source data.
Nitrogen deprivation of yeast enhanced eIF5A H (Hyp2 in yeast) in a Spe1-dependent fashion (Fig. 6b,c and Extended Data Fig. 9a,b ). The hypusination defect observed in Spe1-deficient cells was reversed by SPD (Extended Data Fig. 9c,d ), which had no additional impact on hypusine levels in starved WT cells (Extended Data Fig. 9e,f ).
Similarly, immunostaining of brains revealed increased eIF5A H and total eIF5A in female and male flies after 12 h fasting (Fig. 6d,e and Extended Data Fig. 9g,h ), as confirmed by immunoblotting of whole-head lysates from female flies (Extended Data Fig. 9i,j ). Notably, female heterozygous Odc1 MI10996 mutants, which failed to show lifespan extension upon IF 12:12 , also lost their eIF5A H fasting response (Extended Data Fig. 9k,l ).
We also found elevated eIF5A H in lysates from fasted worms (Fig. 6f,g ) and livers from fasted male mice (Fig. 6h,i ). Of note, dnPI3K hearts, which had increased SPD levels, showed highly upregulated eIF5A H levels, while the IGF1R tg mutation caused no alterations in eIF5A H or SPD levels (Extended Data Fig. 9m,n ).
Increased eIF5A H , but not total eIF5A, levels were similarly detected in starved human U2OS cells (Extended Data Fig. 9o,p ), as well as in PBMCs from healthy human volunteers over several days of fasting, which persisted after food re-introduction (Fig. 6j ). Fasting enhanced eIF5A H indistinguishably in male and female volunteers, and this effect was not affected by age, BMI or weight loss (Extended Data Fig. 9q–t ).
We next explored the mechanisms of increased eIF5A H levels. In yeast, protein levels of Dys1 (DHS) tendentially, but not significantly ( P = 0.077), increased after 6 h and decreased after 24 h nitrogen deprivation, whereas Lia1 (yeast DOHH) was elevated significantly at both time points (Fig. 6k,l and Supplementary Fig. 5a,b ), suggesting a superior role for DOHH in driving or maintaining the observed effects on eIF5A H . Compared with WT cells, ∆ spe1 cells exhibited increased Lia1 protein levels in control but not −N conditions, and this phenotype could be corrected by SPD (Supplementary Fig. 5c,d ). In double knockout ∆ spe1 ∆ lia1 cells, SPD failed to enhance eIF5A H levels in control and −N medium (Supplementary Fig. 5e,f ), indicating that Lia1 is indeed responsible for the hypusination response.
Similarly, increased mRNA transcript levels of dohh-1 ( C. elegans DOHH) and dhps-1 (DHS), but not iff-1 ( EIF5A ) were found in worms after 24 h fasting (Fig. 6m ). Only DOHH levels stayed elevated after prolonged 48 h fasting (Supplementary Fig. 5g ). In U2OS cells, 6 h starvation increased the abundance of mRNAs transcribed from all three genes (Fig. 6n ).
Next, we compared the proteomic landscape during nitrogen deprivation in yeast cells treated with GC7, a specific pharmacological inhibitor of DHS/DYS1, and/or lacking SPE1 . As previously observed, proteins involved in autophagy, TOR signalling, translation and amino acid metabolism were dysregulated when polyamine synthesis or hypusination were suppressed (Supplementary Fig. 5h–k ), implying a multipronged effect of intracellular polyamine metabolism on these processes. Targeting hypusination directly with GC7 phenocopied many of the proteomic perturbations observed in the ∆ spe1 strain, supporting a role of eIF5A H as a major downstream mediator of polyamine effects on energy and amino acid metabolism, as well as on TOR signalling and autophagy (Supplementary Fig. 5h–k ). Additionally, ribosomal processes, which are known to require polyamines 55 , 56 , were strongly decreased when SPE1 was lacking and/or eIF5A H was inhibited by GC7 (Supplementary Fig. 5h,j ). Accordingly, deleting the starvation-responsive LIA1 gene significantly decreased the long-term survival benefits conferred by nitrogen deprivation (Fig. 6o ). Similarly, a temperature-sensitive mutation of eIF5A ( hyp2-1 ) entirely abolished the beneficial effect of nitrogen deprivation on chronological lifespan at the restrictive temperature (28 °C), when eIF5A H was blocked (Supplementary Fig. 6a,b ), but only partially at the permissive temperature (20 °C) (Supplementary Fig. 6c ). Moreover, GC7 reduced eIF5A H (Supplementary Fig. 6d,e ) and chronological lifespan extension by nitrogen depletion (Supplementary Fig. 6f,g ).
A heterozygous point mutation in eIF5A mutating lysine 51, which is the target of hypusination in flies ( eIF5A K51R ), has previously been shown to render SPD supplementation ineffective on the climbing ability of aging flies 53 . Likewise, IF 12:12 conferred lifespan extension was lost in female and male eIF5A K51R /+ flies (Supplementary Fig. 6h,i ), while not affecting food consumption during the first cycles of IF 12:12 (Supplementary Fig. 6j ). Similarly to SPD supplementation 53 , IF 12:12 did not improve the climbing ability of aged female eIF5A K51R /+ flies (Supplementary Fig. 6k–m ).
In C. elegans , the knockdown of dhps-1 (Extended Data Fig. 10a ) attenuated the induction of autophagy (Extended Data Fig. 10b–g ) and strongly reduced the beneficial effect of IF 48:48 on lifespan (Fig. 6p ) without affecting body size (Extended Data Fig. 10h ). Similarly, GC7 reduced autophagic flux in starved U2OS cells (Extended Data Fig. 10i–k ).
In summary, fasting induced SPD-dependent increases in eIF5A H in multiple species and this effect on eIF5A H was required for the pro-longevity effects of nitrogen starvation or IF in yeast, worms and flies.
Acute fasting stimulates autophagy by inhibiting nutrient sensors (such as TORC1 and EP300) and activating signal transducers for nutrient scarcity (such as AMPK and SIRT1) 4 , 57 . Adult-onset IF with CR represents a translatable tool to improve age-associated diseases and systemic health of humans. Still, mechanistic details into the molecular and metabolic relay of nutritional information to lifespan regulation are missing but mandatory for successful clinical implementation. Here, we focused on closing a major gap in our understanding of the metabolic control of fasting-induced autophagy and longevity that apparently involves an increase in SPD-dependent hypusination of eIF5A.
Although fasting reduces the levels of amino acids, including ARG (together with its product ORN) and MET (and its product SAM), it also stimulates metabolic flux through the polyamine synthesis pathway, favouring an increase in SPD levels across different species, which requires ODC1. SPD is needed to hypusinate and activate eIF5A, a translation factor known to stimulate autophagy 58 . Indeed, we detected fasting-induced eIF5A hypusination across all analysed species, supporting the concept that SPD is universally implicated in the fasting response. Accordingly, genetic or pharmacological inhibition of SPD elevation or eIF5A hypusination curbed autophagy induction by fasting, and the longevity-promoting, cardioprotective and antiarthritic effects of IF across the phylogenetic spectrum.
Among the three ODC1-dependent polyamines, SPD seems to be crucial for mediating many fasting responses, mainly because it represents the sole co-factor for eIF5A hypusination. Nevertheless, supplementation with PUT or SPM also reversed ODC1 deficiencies, likely because they are interconvertible with SPD. However, PUT and SPM may partly function by other, yet-to-be-identified mechanisms.
We also found that autophagy induction by pharmacological mTOR inhibition partly depends on SPD synthesis in yeast and human cells, arguing in favour of a general role of SPD in autophagy stimulation. Notably, in yeast, nitrogen deprivation-induced inhibition of TORC1 was partially delayed in ∆ spe1 cells, indicating a reciprocal relationship between polyamines and TOR signalling. Reflecting a complex crosstalk between polyamines and mTOR, SPD treatment was previously shown to activate mTORC1 in the white adipose tissue (WAT) of young mice, but to inhibit mTORC1 in the WAT of aged mice fed a high-fat diet, and not to affect liver mTOR signalling 59 , suggesting organ- or cell type-specific circuitries that remain to be explored.
Of note, in long-lived dilp2-3,5 mutant flies (which lack three of the seven insulin-like peptides), autophagy is induced as a result of enhanced levels of glycine N -methyltransferase (GNMT), which also results in enhanced synthesis of the pro-autophagic metabolite SPD 60 . GNMT overexpression is sufficient to increase lifespan in flies, and lifespan extension via dietary restriction partially depends on GNMT in flies 61 . Similarly, ODC-1 is highly upregulated in the well-studied long-lived C. elegans daf-2 mutant 62 , which encodes for the IGF1 receptor 63 , 64 , further suggesting that the polyamine and insulin signalling pathways are intertwined. Moreover, the liver-specific knockout of insulin receptor substrate 1 (IRS1) causes an increase in GNMT levels in mice 60 , pointing to a phylogenetically conserved pathway linking reduced trophic signalling to SPD elevation. However, it remains to be determined whether this pathway is active in response to fasting. Recently, the longevity effects of IF were linked to the circadian regulation of autophagy in flies 65 . As polyamines are subject to, and regulate, circadian rhythms 66 , this adds yet another possible intersection of lifespan regulation by SPD and hypusination in the context of fasting regimes.
The exact kinetics of the polyamine-eIF5A H response to IF or CR remains unclear on a cell- and tissue-specific level. In future long-term studies, it will be important to weigh the impact of fasting length/periodicity versus the level of CR on polyamine metabolism. In rodents and humans, isocaloric IF studies have produced mixed results on health outcomes and lifespan and it remains to be studied how polyamines mechanistically integrate into this complexity. Given the observation that serum SPD levels were increased in mice after eight months of CR, it will be important to determine how pharmacological or genetic inhibition of polyamine metabolism affects the extension of healthspan and lifespan conferred by CR rather than by IF. While our work focused on the effects downstream of polyamine metabolism, additional studies should dissect the molecular relays connecting nutrient status to the transcriptional, translational and post-translational regulation of polyamine metabolism and spermidine-dependent eIF5A hypusination. Currently, the biochemical mechanism through which IF and CR stimulate polyamine synthesis and subsequent eIF5A hypusination remain elusive, limiting the novelty of our study. Thus, future work must elucidate the crosstalk between polyamines and nutrient-responsive factors, such as insulin/IGF1 and mTOR that inhibit autophagy and accelerate aging or AMPK that induces autophagy and favourably influences healthspan and lifespan. It will also be important to determine whether polyamines induced by IF always confer health benefits. While oral spermidine supplementation can elicit anticancer effects 67 , 68 , elevated polyamine levels are detected in many cancer types and may stimulate cellular proliferation 69 . Thus, the impact of IF on patients with cancer remains to be carefully evaluated.
Our study reveals that fasting-induced longevity and improved healthspan partially rely on SPD-dependent eIF5A hypusination and ensuing autophagy induction in multiple species.
All experiments were conducted in compliance with relevant ethical regulations, as indicated in the respective sections.
BY4741 ( MATa his3 Δ 1 leu2 Δ 0 met15 Δ 0 ura3 Δ 0 ) was used for all experiments, except for experiments with rapamycin in which we used BY4742 ( MATα his3 Δ 1 leu2 Δ 0 lys2 Δ 0 ura3 Δ 0 ). The null mutants BY4741 ∆ spe2 ::kanMX ( Y01743 ), ∆ spe3 ::kanMX ( Y05488 ), ∆ spe4 ::kanMX ( Y06945 ) and the temperature-sensitive mutant hyp2-1 :kanMX (TSA736) were obtained from Euroscarf. We generated the null mutants BY4741 ∆ spe1:: hphNT1, ∆ lia1 ::natNT2 and ∆ spe1:: hphNT1∆ lia1 ::natNT2 according to previously described methods 70 . In brief, we amplified a gene-specific hphNT1 or natNT2 knockout cassette by PCR using the template plasmids pFA6a-hphNT1 and pFA6a-natNT2 (ref. 70 ) and primers:
SPE1_pYM_S1: 5′-GTTCTACAACTTTTTCATAGTAATCAAAACCTTTGAATTTCAAACTTACTATGCGTACGCTGCAGGTCGAC-3′
SPE1_pYM_S2: 5′-CTTTTCCCACCCCCTCCGTCTCTCTTGCGAAAGTCGTGGTTAAATATATCCTTCAATCGATGAATTCGAGCTCG-3′
Forward control primer SPE1_Ctrl: 5′-TCATCAAGAGCCCCATCC-3′
Reverse control primer Control_S1: 5′-GTCGACCTGCAGCGTACG-3′
LIA1_pYM_S1: 5′-GTTAGGATAAACTGTAGTCCTTCTAACATACCACGCAAGAAAGAAAAAAAAAAACCGTACGCTGCAGGTCGAC-3′
LIA1_pYM_S2: 5′-CAAGATTATACAATGATTATTGTTACTATCATTATTGATGATGCTGATTCTTATCGATGAATTCGAGCTCG-3′
Forward control primer LIA1_Ctrl: 5′-GTTCCCAGGCGAAGAAAGAAC-3′
Reverse control primer natNT2_Ctrl: 5′-CGTGTCGTCAAGAGTGGTAC-3′
To determine the protein levels of Dys1 and Lia1, we generated genome-tagged C-terminal 6×HA fusions using the plasmid pYM16 (ref. 70 ) and the following primers:
For Dys1-6×HA:
DYS1_pYM_S3: 5′-CTGCTACCTTTGCCAGTGGTAAACCAATCAAAAAAGTTAAGAATCGTACGCTGCAGGTCGAC-3′
DYS1_pYM_S2: 5′-GAAATAGTACAGATTCATTTTTTTTTTTTTCATCTCAAAATTCTCTCATCAATCGATGAATTCGAGCTCG-3′
Forward control primer DYS1_Ctrl: 5′-GCGTGACCAAGGTATGAATCGTATT-3′
Reverse control primer Control_hphNT1: 5′-CATATCCACGCCCTCCTAC -3′
For Lia1-6×HA:
LIA1_pYM_S3: 5′-GATATGTATGATTACGAAAACAGCAACGAACTAGAATATGCTCCAACTGCTAATCGTACGCTGCAGGTCGAC-3′
LIA1_pYM_S2: 5′-CAAGATTATACAATGATTATTGTTACTATCATTATTGATGATGCTGATTCTTCTAATCGATGAATTCGAGCTCG-3′
Forward control primer LIA1_Ctrl: 5′-CTAGGTGACAAGGATTCGTTGGATG-3′
Reverse control primer Control_hphNT1: 5′- CATATCCACGCCCTCCTAC-3′
For the Pho8∆N60 assay to measure nonselective autophagy, we generated PHO8 pho8∆N60-URA3 strains in BY4741 WT and ∆ spe1 background (in this case, the ∆ spe1 strain was obtained from Euroscarf ; spe1 ::kanMX; Y05034 ) using the integrative pTN9-URA plasmid 71 . Pho8∆N60-specific alkaline phosphatase activity was determined as described previously 72 . Endogenous GFP–Atg8 fusions were generated as previously described 72 . In brief, we used the vector template pYM-pATG8 (ref. 72 ) to construct NatNT2::P ATG8 -yeGFP-ATG8 strains with primers:
Atg8_pYM_S1: 5′-CTAATAATTGTAAAGTTGAGAAAATCATAATAAAATAATTACTAGAGACATGCGTACGCTGCAGGTCGAC-3′
Atg8_pYM_S4: 5′-GACTCCGCCTTCCTTTTTTCAAATGGATATTCAGACTTAAATGTAGACTTCATCGATGAATTCTCTGTCG-3′
Forward control primer yeGFP_F: 5′-GGTGAAGAATTATTCACTGGTGTTG-3′
Reverse control primer ATG8_R: 5′-GAACAATAGATGGCTAATGAGTCC-3′
Medium and growth conditions
All experiments were performed in baffled 100-ml flasks filled with 10–15 ml growth medium, incubated at 28 °C and constant shaking (145 rpm), if not stated otherwise. For general experiments, we used standard dextrose medium (CTL) containing 0.17% yeast nitrogen base (BD Diagnostics), 0.5% (NH 4 ) 2 SO 4 , 80 mg l −1 histidine, 200 mg l −1 leucine 320 mg l −1 uracil, 30 mg l −1 adenine and 30 mg l −1 all other amino acids, with 2% glucose, which we sterilized as separate 10× stocks by autoclaving. For experiments with BY4742 we additionally added 90 mg l −1 lysine after mixing the stocks.
All cultures were inoculated from cellular material from a YPD (1% yeast extract (BD), 2% bacto peptone (BD), 2% glucose, 2% agar) plate, which was incubated at 28 °C for 2–3 days, placed at 4 °C for at least 1 day and a maximum of 2 weeks. We used pre-cultures in CTL medium to inoculate main cultures to an OD 600 of ~0.01–0.05. In general, main cultures were grown to mid-logarithmic phase (OD 600 = 1), centrifuged at 3,000–4,000 rpm (1731–3077 rcf) for 3–5 min, washed once with sterile pre-warmed ddH 2 O, and re-suspended in fresh medium. For nitrogen deprivation (−N) we re-suspended the cells in fresh CTL or −N medium, containing 0.17% yeast nitrogen base and 2% glucose. For glucose restriction, we used CTL medium with either 0.5% or 0.05% glucose. For water starvation, we used sterile ddH 2 O.
Rapamycin and GC7 treatments
Rapamycin (LC Laboratories, R-5000) stock was prepared as 1.1 mM in dimethylsulfoxide (DMSO) (stored at −20 °C) and used at a final concentration of 40 nM. Analogous to the −N experiments, we initiated rapamycin treatment by adding DMSO as a control or rapamycin to the cultures during the mid-logarithmic phase (at OD 600 = 1). The pharmacological DHS inhibitor N1-guanyl-1,7-diamine-heptane (GC7) was purchased from Merck (259545) and prepared as a sterile filtered 50 mM stock in ddH 2 O, stored at −20 °C. GC7 treatment was initiated during inoculation and repeated when the medium was changed for nitrogen deprivation.
Yeast polyamine supplementation
Generally, polyamines were supplemented from sterile stocks of polyamines starting with the main culture, except for synergy experiments as shown in Fig. 3k,l , Supplementary Fig. 4c and Extended Data Figs. 4m,n and 9e,f , where spermidine was added in the starvation medium only. For the other nitrogen starvation experiments, polyamine supplementation was renewed in the fresh medium. Cells were treated with either 100 µM putrescine (Sigma, D13208; from 3 M aqueous stock, pH 7.4), spermidine (Sigma, 85558; from 1 M aqueous stock, pH 7.4) or spermine (Sigma, 85590; from 0.9 M aqueous stock, pH 7.4) or 5 mM spermidine as indicated in the figure legends. In brief, the polyamine stocks were prepared on ice, with ddH 2 O and the pH was set with HCl, as previously described 23 . Stock solutions were stored at −20 °C for up to one month.
Yeast autophagy measurements
To monitor yeast autophagy, we employed established protocols using fluorescence microscopy, western blots and biochemical assays 73 . We used endogenously tagged eGFP–Atg8 fusion strains, as previously reported 72 .
GFP–Atg8 localization was assessed by collecting 100 µl from yeast cultures by centrifugation (1,000 rcf, 1 min) and re-suspension of the cell pellet in 50 µl propidium iodide (PI) staining solution (100 ng ml −1 PI in PBS, pH 7.4). Cells were pelleted again (500 rcf, 30 s), transferred to a glass microscopy slide, and covered with a cover slip. Imaging was performed with a Leica DM6B-Z fluorescence microscope using a ×100/1.40 HC Pl APO oil objective and a Leica-DFC9000GT-VSC09095 sCMOS camera. eGFP images were taken with a GFP filter (Ex 470 /Em 525 ) filter and 2 s exposure. PI images were taken with a Texas Red filter (Ex 560 /Em 630 ) and 75-ms exposure. Raw 16-bit images were exported and merged with constant contrast settings (eGFP 3,000–30,000; PI 200–50,000) in ImageJ 74 . Cells were classified manually in a blinded fashion using the ImageJ CellCounter tool.
Additionally, we performed Pho8∆N60 assays 43 , as previously reported 72 . In brief, we collected three OD 600 equivalents at the indicated time points by full-speed centrifugation in a standard tabletop centrifuge for 2 min. After a washing step with ddH 2 O, we re-suspended the cell pellet in 200 µl cold assay buffer (250 mM Tris-HCl (pH 9), 10 mM MgSO 4 and 10 μM ZnSO 4 ) and transferred it to pre-cooled plastic reaction tubes filled with 100 µl acid-washed glass beads. The cells were homogenized in a BeadBeater (2 × 1 min with 1-min pause between the cycles) in a liquid nitrogen-cooled metal rack. After homogenization we centrifuged the samples at 10,000 rcf for 10 min at 4 °C and 100 µl supernatant was transferred into a fresh pre-cooled tube. We used a Bio-Rad protein assay (Bio-Rad, 5000006) to determine the protein concentration of the supernatant. Protein extract corresponding to 1.5 µg protein was transferred to a well of a 96-deep-well plate in duplicate and filled with assay buffer to 550 µl at room temperature (22–25 °C). The addition of 50 µl α-naphthyl phosphate solution (55 mM in assay buffer, pH 9) started the reaction. After mixing the plate by vortexing, it was incubated for 20 min at 30 °C. The reaction was stopped by adding 200 μl stopping buffer (2 M glycine/NaOH, pH 11) to each well, followed by vortexing. Then, 100 µl of each well was transferred to a 96-well plate (black bottom) in duplicate and measured in a TECAN plate reader (Ex 340 and Em 485 ). For the correction of background phosphatase activity, control strains (without the Pho8∆N60 mutation) were processed in parallel and subtracted.
Yeast immunoblotting
A culture volume corresponding to three OD 600 units was collected at each time point by centrifugation at 3,000 rcf for 3 min. After a washing step with ddH 2 O, the cell pellet was frozen at −20 °C until further processing. Whole-cell extracts were generated by re-suspending the cell pellets in 300 µl lysis buffer (1.85 M NaOH and 7.5% 2-mercaptoethanol) and incubation on ice for 10 min. Proteins were precipitated by adding 300 µl 55% trichloroacetic acid (TCA), incubation on ice for 10 min, centrifugation at 10,000 rcf at 4 °C for 10 min and removal of the supernatant. After an additional centrifugation step (10,000 rcf at 4 °C for 2 min) residual supernatant was removed and pellets were solubilized in final sample buffer (62.5 mM Tris-HCl, pH 6.8, 2% SDS, 8.7% glycerol, 0.004% bromophenol blue and 120 mM dithiothreitol (DTT)) and 1 M Tris was added to neutralize residual TCA until the samples turned blue. In brief, before electrophoresis, samples were boiled at 95 °C for 5 min and centrifuged at maximum speed in a tabletop centrifuge for 15 s. For protein separation, 12–15 µl of the supernatant was loaded on 4–12% or 12% (for GFP–Atg8) NuPAGE Bis-Tris gels (Thermo Fisher Scientific). Electrophoresis was performed at 100–130 V with MOPS SDS running buffer (Thermo Fisher Scientific, NP000102). Proteins were wet-transferred to methanol-activated 0.45-µm PVDF membranes (Roth, T830.1) at 220 mA for 60–90 min using transfer buffer (10 mM CAPS/NaOH, pH 11 and 10% methanol). After blotting, membranes were blocked with blocking solution (1% dry milk powder in Tris-buffered saline (TBS), pH 7.4) for 1 h and then incubated with the primary antibodies overnight at 4 °C. After three washing steps in TBS + 0.1% Triton X-100 for 5 min, membranes were incubated with secondary, horse radish peroxidase (HRP)-linked antibodies for 1 h at room temperature. After three washing steps in TBS + 0.1% Triton X-100 for 5 min, signals were detected with a ChemiDoc detection system (Bio-Rad) and Clarity Western ECL Substrate (Bio-Rad) using the ‘optimal exposure’ setting. For re-probing membranes, Restore PLUS Western Blot Stripping Buffer (Thermo Fisher Scientific, 46430) was used according to the manufacturer’s protocol. Band intensities were quantified using ImageLab v.5.2 (Bio-Rad) using the rectangular volume tool with local background adjustment. Primary antibodies were anti-GFP (Roche, 1814460, 1:5,000 dilution in blocking solution), anti-hypusine (Merck, ABS1064-I, 1:1,000 dilution in blocking solution), anti-GAPDH clone GA1R (Thermo Fisher Scientific, MA5-15738, 1:10,000 dilution in blocking solution) and anti-HA (Sigma, H-9658, 1:10,000 dilution in blocking solution). Secondary antibodies were HRP-linked anti-mouse IgG (Sigma, A9044, 1:10,000 in blocking solution) or HRP-linked anti-rabbit IgG (Sigma, A0545, 1:10,000 dilution in blocking solution).
Yeast chronological lifespan
Chronological aging experiments were performed as previously described 75 . In brief, we used PI staining to identify dead cells using an LSR II Fortessa flow cytometer equipped with a high-throughput sampler (BD), using FACSDiva software (BD). For that purpose, 20-µl aliquots of the cultures were transferred to 96-well U-bottom plates at the indicated time points and stained with 150 µl PI staining solution (100 ng ml −1 in PBS), incubated in the dark for 15 min at room temperature and centrifuged at 2,500 rcf for 5 min. The staining solution was removed by tapping the plate and the cells were re-suspended in 150 µl fresh PBS. Then, 30,000 cells were measured per sample and PI-positive (dead)/negative (live) cells were identified and quantified by flow cytometry at Ex 488 /Em 670 .
At selected time points, we additionally performed live qualitative fluorescence microscopy, for which we took 100-µl aliquots of the cultures, centrifuged them for 1 min at room temperature in a standard tabletop centrifuge and re-suspended the pellet in 100 µl PI staining solution. After 2 min of incubation in the dark, we transferred a droplet of this suspension onto glass microscopy slides and covered them with coverslips. Images were taken with a Leica DM6B-Z microscope, using the Texas Red filter (Ex 560 /Em630, 300-ms exposure). For image processing, we used ImageJ 74 .
Of note, yeast chronological lifespan is affected by the medium pH 76 , which was measured with a Thermo Scientific Orion Star A221 pH meter and only minimally affected during nitrogen starvation by the SPE1 knockout (Supplementary Fig. 4a ).
Yeast replicative lifespan
Yeast replicative lifespan was performed as previously described 77 , 78 . To summarize, yeast was pulled from −80 °C freezer stocks onto YPD plates and were allowed to grow at 30 °C for 2 days. Single colonies from each sample were picked and patched for two consecutive days before being plated to RLS assay plates (YPD (1% yeast extract, 2% peptone and 2%, 0.5% or 0.05% glucose) unless otherwise stated). Virgin cells were selected as new mother cells and each generation of daughter cells were separated and counted via microdissection. Cell death is considered when mother cells are no longer capable of dividing. Mother cells that divided two or fewer times or were lost during the experiment were censored from the dataset.
Yeast TORC1 activity
In vivo TORC1 activity was assayed as previously described 79 . In brief, a 10-ml cell culture was mixed with TCA at a final concentration of 6%. After centrifugation, the pellet was washed with cold acetone and dried in a SpeedVac. The pellet was re-suspended in lysis buffer (50 mM Tris-HCl, pH 7.5, 5 mM EDTA, 6 M urea and 1% SDS), the amount being proportional to the OD 600 of the original cell culture. Proteins were extracted by mechanical disruption in a Precellys machine after the addition of glass beads. Subsequently, an amount of 2× sample buffer (350 mM Tris-HCl (pH 6.8), 30% glycerol, 600 mM DTT, 10% SDS and BBF) was added to the whole-cell extract and the mix was boiled at 98 °C for 5 min. The analysis was carried out by SDS–PAGE by loading 15 µl extracts on 7.5% polyacrylamide gels. The transfer on nitrocellulose membranes was performed with a Trans-Blot Turbo Transfer System (Bio-Rad) with Bjerrum Schafer-Nielsen buffer (48 mM Tris-HCl (pH 9.2), 39 mM glycine and 20% methanol). Protein detection was carried out using custom phosphospecific rabbit anti-Sch9-pThr737 (1:10,000 dilution in TBS 1% milk) and goat anti-Sch9 (1:1,000 dilution in TBS 5% milk) antibodies and a commercial rabbit anti-Adh1 antibody (Calbiochem, 126745, 1:50,000 dilution in TBS 5% milk). Band intensities from independent biological replicates were quantified with ImageJ.
Yeast metabolomics
Cells (equivalent to 15 OD 600 , grown as described before) of BY4741 and BY4741 ∆ spe1 after 6 or 24 h of nitrogen deprivation were collected by centrifugation at 4,000 rpm (3,077 rcf) for 3 min at 4 °C, washed once with ice-cold ddH 2 O, frozen in liquid nitrogen and stored at −80 °C until further processing. The pellets were re-suspended in 400 µl ice-cold methanol (−20 °C) and 200 µl MilliQ H 2 O and transferred to a tube containing Precellys beads (1.4-mm zirconium oxide beads, Bertin Technologies, Villeurbanne, France) for homogenization on a Precellys 24 homogenizer for two cycles of 20 s with 5,000 rpm and 10 s breaks. Cell debris was pelleted by centrifugation at 13,000 rpm (15,871 rcf) for 30 min (4 °C). Supernatants were lyophilized and dissolved in 500 µl NMR buffer (0.08 M Na 2 HPO 4 , 5 mM 3-(trimethylsilyl) propionic acid-2,2,3,3-d 4 sodium salt (TSP), 0.04% ( w / v ) NaN 3 in D 2 O, pH adjusted to 7.4 with 8 M HCl and 5 M NaOH) for NMR measurements.
All NMR experiments were acquired at 310 K using a Bruker 600 MHz Avance Neo spectrometer equipped with a TXI probe head. The one-dimensional (1D) CPMG (Carr–Purcell–Meiboom–Gill) pulse sequence (‘cpmgpr1d’; 512 scans; size of free induction decay (FID) 73,728; 11,904.76 Hz spectral width; recycle delay 4 s), with water signal suppression using presaturation, was recorded for 1 H 1D NMR experiments. In brief, and as described before 80 , data were processed in Bruker Topspin v.4.0.6 using one-dimensional exponential window multiplication of the FID, Fourier transformation and phase correction. The NMR data were then imported into Matlab2014b; TSP was used as the internal standard for chemical-shift referencing (set to 0 ppm); regions around the water, TSP and methanol signals were excluded; the NMR spectra were aligned; and a probabilistic quotient normalization was performed. The statistical significance of the determined differences was validated by the quality assessment statistic Q 2 . This measure provides information about cross-validation and is a qualitative measure of consistency between the predicted and original data with a maximum value of 1.
Metabolite identification was carried out using Chenomx NMR Suite v.8.4 (Chenomx) and reference compounds. Quantification of metabolites was carried out by signal integration of normalized spectra. For each metabolite, a representative peak with no overlapping signals was identified, the start and end points of the integration were chosen to revolve around that peak, and the area of the peak was integrated by summing up the value for each point. Peaks which could not be unambiguously identified are shown with their ppm value at maximal intensity.
We used MetaboAnalyst v.5.0 for data analysis and visualization 81 , using mean-centred normalization. For metabolite set enrichment analysis we used the integrated KEGG database. sPLS-DA analysis was performed with ten features for each component and heatmaps were generated using Ward’s method and Euclidian distance settings.
Yeast polyamine measurements
Polyamine extraction and quantification was performed as previously described 82 . In brief, yeast cells were grown as previously mentioned and at the indicated time points, one to three OD 600 were collected by centrifugation, washed once with ddH 2 O and mixed with 5% TCA containing stable-isotope labelled standards of indicated polyamines at a final concentration of 100 ng ml −1 each, followed by incubation on ice for 60 min with vortexing every 15 min. After centrifugation at 25,000 rcf at 4 °C for 10 min, 150 µl supernatant were transferred to 1.5-ml LoBind reaction tubes and 37.5 µl ammonium formate (2 M), 800 µl of ultra-pure H 2 O and 125 µl of saturated Na 2 CO 3 buffer were added.
For derivatization, 20 µl isobutyl chloroformate was added, followed by a 15-min incubation at 35 °C. After centrifugation for 1 min at 15,000 rpm/21,130 rcf, 800 µl supernatant was then transferred to LoBind 96-deep-well plates (VWR International, 737-2544) for storage at −80 °C. Polyamine derivatives were extracted offline by solid-phase extraction (SPE) (Strata-X, Polymeric Reversed Phase, 96-well plate). SPE was conditioned with 500 µl acetonitrile, equilibrated with 500 µl distilled water containing 0.2% acetic acid. Derivatized TCA extracts were loaded onto the SPE and after two washing steps with 500 µl 0.2% acetic acid, samples were eluted with 250 µl 80% acetonitrile containing 0.2% acetic acid. Eluted SPE extracts were subjected to liquid chromatography (LC)–MS/MS (mobile phase, isocratic 80% acetonitrile containing 0.2% acetic acid; flow rate 250 µl min −1 ; HPLC column: Kinetex 2.6 µm C18 100A 50 mm × 2.1 mm TSQ Quantum Access Max coupled to an Ultimate 3000). MS conditions were set as previously published 83 . LC–MS/MS data were acquired and processed using Xcalibur v.4.0 Software (Thermo Fisher Scientific). The final data were normalized to OD 600 and the control conditions for each polyamine.
Yeast arginine flux metabolomics
The main cultures were grown to log phase in medium containing 13 C 6 -labelled arginine (30 mg l −1 ; Eurisotop, CLM-2265) instead of non-labelled arginine. The control cells received fresh medium containing 13 C 6 -labelled arginine, whereas the nitrogen-starved cells were cultivated in standard −N medium for another 6 h, before collecting by centrifugation as described above. Metabolites were measured following a previously described protocol 84 , but without internal standards in the extraction buffer. In brief, 50-µl samples were vortexed for 5 min with 500 µl ice-cold extraction mixture (methanol:water, 9:1, −20 °C) and then centrifuged (10 min at 15,000 rpm/21,130 rcf, 4 °C). Several fractions were then split to be analysed by LC–MS and gas chromatography (GC)–MS 84 . One fraction was analysed by the LC–MS Orbitrap q-Exactive Thermo in full profiling. Tracking of arginine and its isotopes was performed by post-acquisition treatment with the software Xcalibur Thermo (v.2.2). Polyamines analyses were performed by LC–MS/MS with a 1290 UHPLC (ultra-high performance liquid chromatography) (Agilent Technologies) coupled to a QQQ 6470 (Agilent Technologies) and were previously described 85 . The MRM method was adapted to track polyamines but also their isotopes. Data were processed by MassHunter Quantifier (Agilent) software (v.10.1).
Yeast proteomics
Yeast samples were processed in each sequence (experiment) in one MS run to reduce batch effects. For cell lysis 5% SDS, 50 mM ammonium bicarbonate (used in the GC7-pertubation experiment) or 8 M urea, 20 mM HEPES (used in the SPD-rescue experiment) and zirconia beads (1:1 v / v ) were added to the frozen pellet and lysed in a BeadBeater in three cycles (2 min, 15 s rest, 1,500 rpm per cycle) (Spex Geno Grinder). Then, 150 µg protein of each sample were alkylated with 5 mM tris(2-carboxyethyl)phosphine (TCEP), reduced with 10 mM chloroacetamide (CAA) and digested with trypsin (1:50 wt / wt ) as well as purified utilizing the S-Trap mini columns (Protifi), following the manufacturer’s instructions: S-Trap mini spin column digestion protocol. Peptides were lyophilized, re-suspended in 0.1% formic acid and diluted to 200 ng µl −1 , whereby 1 µl was used per MS injection.
Samples were analysed on a timsTOF ion mobility mass spectrometer (with PASEF technology, Bruker Daltonics) in-line with UltiMate 3000 RSLCnano UHPLC system (Thermo Scientific). Peptides were separated on a reversed-phase C18 Aurora column (25 cm × 75 µm) with an integrated Captive Spray Emitter (IonOpticks). Mobile phases A 0.1% ( v / v ) formic acid in water and B 0.1% ( v / v ) formic acid in ACN with a flow rate of 300 nl min −1 , respectively. Fraction B was linearly increased from 2% to 25% in the 90-min gradient, increased to 40% for 10 min, and a further increase to 80% for 10 min, followed by re-equilibration. The spectra were recorded in data-independent acquisition (DIA) mode as previously described 86 .
The DIA data were quantified with DIA-NN v.1.8.0 (ref. 87 ) by creating a synthetic fasta library using the reviewed Uni-Prot S. cerevisiae protein database (downloaded 24 September 2021). The MS proteomics raw data of both experiments together with the processing log files have been deposited to the ProteomeXchange Consortium using the PRIDE partner repository ( https://www.ebi.ac.uk/pride/ ) with the dataset identifier PXD035909 . In the SPD-rescue experiment, a total of 4,684 distinct protein groups were identified. Notably, 82% of these groups demonstrated consistent presence across all samples, indicating a robust and pervasive occurrence. Furthermore, close to 90% of the samples exhibited the presence of approximately 4,207 protein groups, corroborating their widespread and persistent detection within the experimental set. In the GC7-perturbation experiment, 4,019 unique protein groups were quantified. Among these, 45% were identified across every sample, with approximately 58% detected in at least 90% of the samples.
PCA was performed by singular-value decomposition of the centred and scaled protein groups ( n = 4,684), whereby missing values were imputed with zero. Protein complex enrichment analyses were performed based on KEGG yeast pathways from ConsensusPathDB (downloaded 27 April 2022) 88 , as well as complex portal annotations of the TORC complex 89 . Aggregated z -values were calculated for each protein complex resulting from averaged absolute protein expression of each complex member. The z -scores for each protein were calculated as followed \(z=\frac{\left(x-\mu \right)}{\sigma }\) , where x is the expression, μ is the protein expression across all samples and σ is the s.d. of the protein expression across all samples.
Data were visualized using R 90 by utilizing the packages prcomp 91 , 92 , ggplot 93 and pheatmap 94 .
Fly strains, husbandry and food
All fly strains had a w 1118 genetic background. The heterozygous mutant strains were backcrossed to the laboratory-specific w 1118 at least six times to ensure isogenicity. The heterozygous odc1 mutant was purchased from Bloomington Drosophila Stock center (Indiana University, 56103). The heterozygous eIF5A mutant carrying a point mutation at the lysine site of hypusination was created as described previously 53 . Fly food was prepared according to the Bloomington medium recipe with minor modifications, which we refer to as ‘normal food’ (per litre: 4.2 g agar–agar, 85.3 g sugar beet syrup, 7.5 g baker’s yeast, 8.3 g soybean flour, 66.7 g cornmeal, 1.3 g methyl p-hydroxybenzoate dissolved in 4.2 ml ethanol and 5.25 ml propionic acid). Difluoromethylornithine (DFMO) (a kind gift from P. M. Woster, Medical University of South Carolina), freshly prepared as a sterile filtered 50 mM stock in ddH 2 O, was added to the food after cooling the freshly cooked food to ~40 °C. Agar-only food (0.6%) prepared with a 1:1 mixture of deionized and tap water was used as the fasting food. The flies were reared under standard laboratory conditions, as previously reported (25 °C, 70% humidity with a constant 12-h light–dark cycle) 53 , 95 . Flies used in all experiments were F1. WT flies and mutant flies were collected within 24–48 h after eclosion and considered as 1-day-old flies. The w 1118 virgin females were used to cross with males from w 1118 and mutant strains, respectively. Female and male flies were separated after 24 h of mating on fresh food by light CO 2 anaesthesia and were flipped to fresh normal food every other day in portions of 20–25 flies per vial.
Fly intermittent fasting
IF regimes were initiated 1 day after sorting and maintained throughout the flies’ lifespan. We used a 12-h feeding–fasting cycle (IF 12:12 ), providing food during daylight hours (8:00 to 20:00). Control groups were flipped in the same rhythm but had food access during the whole 24 h. We changed food vials every 2–3 days and kept vials at 4 °C when not used. Flies that escaped or died due to unnatural causes were censored. Dead flies were identified by their unresponsiveness to mechanical stimuli using a small brush.
Fly body weight measurement
Drosophila body weight was measured throughout a fasting cycle as wet weight of snap-frozen fly bodies. The start time of the experiments was 20:00, followed by measurement time points after 12 and 24 h of fasting, as well as after 12 h of re-feeding. The ad libitum-fed group had access to food for the whole time. Data were normalized to each ad libitum group.
Fly food consumption
Food consumption was monitored similarly to the previously described capillary feeding (CAFE) assay 96 : For each replicate, five flies were anaesthetized on ice and transferred to a chamber with two 5-µl glass capillaries (VWR International, 612-2401) inserted in the top caps. Capillaries were either filled with liquid food containing 5% ( w / v ) sucrose (Roth, 4661.4) and 5% ( w / v ) yeast extract (BD, 288620) (ad libitum) or tap water (fasting). Data were collected twice per day (8:00 and 20:00) for at least 3 days after a 24-h acclimatization period. IF flies had access to capillaries containing food from 8:00 to 20:00 Passive evaporation was accounted for with three empty chambers without flies. Data are presented as microlitres of food per fly per hour.
Fly polyamine measurement
Twenty flies (wet weight of snap-frozen fly bodies was estimated and used for normalization) were placed into 1.5-ml LoBind reaction tubes (Eppendorf, 0030108116), snap-frozen in liquid nitrogen and homogenized for 30 s on ice in 600 µl 5% TCA containing stable-isotope labelled standards of indicated polyamines at a final concentration of 100 ng ml −1 each using a Turrax homogenizer (IKA T10 basic). Further polyamine extraction and quantification was performed as previously described with adapted analyte concentrations in the calibration solutions 82 (see also ‘Yeast’ section).
Fly immunoblotting
For fly protein extraction, ten snap-frozen fly heads were homogenized in lysis buffer (1× PBS, pH 7, 0.5% Triton X-100, 2% SDS and 1× complete protease inhibitor mix) for 30 s with a motorized pestle (5 µl lysis buffer per fly head) followed by an incubation for 60 min at 4 °C with continuous rotation. The homogenized samples were centrifuged for 10 min at 10,000 rcf at 4 °C. The final sample buffer (final concentration of 62.5 mM Tris-HCl, pH 6.8, 2% SDS, 8.7% glycerol, 0.004% bromophenol blue and 120 mM DTT) was added to the supernatant and samples were stored at −20 °C until further processing. In brief, before electrophoresis, samples were boiled at 95 °C for 5 min and centrifuged at 10,000 rcf for 1 min. Then, 12.5 µl supernatant (equivalent to two heads) were loaded onto 4–12% Bis-Tris gels and run at 90–120 V. Proteins were transferred to 0.45-μm PVDF membranes for 90 min at 220 mA using transfer buffer (10 mM CAPS pH 11 and 10% methanol). The membranes were then incubated in blocking solution (3% dry milk powder in TBS, pH 7.4) for 1 h and subsequently incubated with the primary antibody overnight at 4 °C. Membranes were washed three times with TBST (TBS + 0.1% Tween-20) for 5 min and subsequently incubated with the secondary antibody for 1 h at room temperature. After three washing steps in TBST for 5 min, signals were detected with a ChemiDoc detection system (Bio-Rad) and Clarity Western ECL Substrate (Bio-Rad) using the ‘optimal exposure’ setting. Band intensities were quantified using ImageLab v.5.2 (Bio-Rad) using the rectangular volume tool with local background adjustment. Primary antibody was anti-hypusine (Merck, ABS1064-I, 1:1,000 dilution in 1% dry milk powder in TBS, pH 7.4). The secondary antibody was HRP-linked anti-rabbit IgG (Sigma, A0545, 1:10,000 dilution in 1% dry milk powder in TBS pH 7.4). As a loading control, HRP anti-β actin monoclonal antibody (Abcam, ab197277, 1:2,000 dilution in TBS + 0.1% Tween +1% BSA) was used.
Fly locomotor function (climbing assay)
For assessing the locomotor function, 5–20 flies were placed in custom-made three-dimensional-printed climbing chambers with nine slots per chamber followed by a defined acclimation period of 30 min in the dark. The chambers were individually attached to a custom-built climbing platform to ensure a standardized impact force and tapped down four times with an interval of 30 s (four rounds). Flies climbing up the rack were recorded by video tracking (Canon EOS 700D). The videos were then analysed with a fly tracking program, which tracks flies based on previously described methods 97 . For the calculation of climbing parameters (height, average height reached by the flies in the given climbing duration; speed, speed of the flies during the climbing duration in mm s −1 ; and distance, the distance travelled by the flies during the climbing duration), flies were tracked over 10 s after the tapping impact. For analysis, the area under the curve (AUC) was calculated for each parameter and strain and normalized to the respective control. Every data point corresponds to the average of the flies assessed in one chamber.
Fly whole-mount immunostaining, confocal imaging and quantification
Flies at 6 days old were either fed ad libitum or fasted for 12 or 24 h. Then the fly brains were dissected for brain immunostaining. Adult brains were dissected in HL3 solution on ice and immediately fixed in cold 4% paraformaldehyde ( w / v ) for 30 min at room temperature. After fixation, samples were washed three times for 10 min each with 0.7% PBT (PBS containing 0.7% Triton X-100 v / v ) and then blocked with 10% normal goat serum in PBT ( v / v ) for 2 h at room temperature. After blocking, samples were incubated in 0.7% PBT containing 5% normal goat serum and primary antibodies for 48 h at 4 °C. After primary antibody incubation, brains were washed in 0.7% PBT six times for 30 min each at room temperature and then incubated in 0.7% PBT with 5% normal goat serum containing the secondary antibodies overnight at 4 °C. Brains were washed six times for 30 min each with PBT at room temperature and mounted in Vectashield (Vector Labs). The following antibodies and dilutions were used in whole-mount adult brain staining: rabbit anti-hypusine antibody (Merck, ABS1064; 1:1,000 dilution), guinea pig anti-eIF5A antibody (customized; 1:200 dilution), goat anti-guinea pig Alexa 555 (Invitrogen; 1:200 dilution), goat anti-rabbit Cy5 (Invitrogen; 1:200 dilution).
Image stacks of specimens were imaged on a Leica TCS SP8 confocal microscope (Leica Microsystems) using a ×40, 1.3 NA oil objective for whole-brain imaging with a voxel size of 0.3788 × 0.3788 × 0.9997 micron 3 . Images were quantified using ImageJ software. In brief, the average intensity z-projection was performed with ~25 stacks of each brain from the beginning of the antennal lobe to the end of the antennal lobe and the mean grey value of the central brain was measured.
C. elegans strains
We followed standard procedures for C. elegans maintenance and other genetic manipulations 98 . The nematode-rearing temperature was kept at 20 °C unless noted otherwise. The following strains, available at the Caenorhabditis Genetics Center, were used: N2: WT Bristol isolate, HZ589: bpIs151 (sqst-1p::sqst-1::GFP + unc-76(+)), MAH215: sqIs11 (lgg-1p::mCherry::GFP::lgg-1 + rol-6).
C. elegans lifespan assays
Lifespan assays were performed at 20 °C. Synchronous animal populations were generated by hypochlorite treatment of gravid adults to obtain tightly synchronized embryos that were allowed to develop into adulthood under appropriate, defined conditions.
For RNAi lifespan experiments, worms were placed on Nematode Growth Medium plates containing 2 mM IPTG and seeded with HT115 (DE3) bacteria transformed with either the pL4440 vector or the test RNAi construct. The progeny was grown at 20 °C unless noted otherwise, through the L4-young adult larval stage and then transferred to fresh plates in groups of around 30 worms per plate for a total of at least 155 individuals per experiment. Animals were transferred to fresh plates every 2 days thereafter and examined every day for touch-provoked movement and pharyngeal pumping until death. Worms that died owing to internally hatched eggs, an extruded gonad or desiccation due to crawling on the edge of the plates were censored and incorporated as such into the dataset. Each survival assay was repeated at least four times.
For IF (IF 48:48 ), synchronized young adult worms raised on Nematode Growth Medium plates with live HT115 were transferred to plates containing floxuridine (120 μM), seeded with HT115 RNAi-transformed bacteria, which were UV-killed before animal transfer. On day 2 of adulthood, worms were divided into ad libitum and IF. Worms in the ad libitum groups were fed ultraviolet-killed HT115 throughout their lifespan. Worms in IF were placed on plates with UV-killed HT115 or without food (seeded bacteria) alternatively every other day. All worms were transferred to freshly seeded plates every 2 days 99 .
C. elegans spermidine and rapamycin treatments
Lifespan assays with spermidine and rapamycin supplementation were performed as previously described 23 , 100 . In brief, SPD (Sigma, S0266) was used at a final concentration of 0.2 mM. Sterilized water solution of SPD was applied to the top of the RNAi bacterial lawn after being killed by UV irradiation for 15 min (0.5 J) using a UV cross-linker (bio-Link BLX-E365;Vilber Lourmat). Rapamycin (LC Laboratories) was dissolved in DMSO and added to the agar plate to a final concentration of 50 μM. Control plates contained an appropriate concentration of DMSO (<0.1%). SPD and rapamycin experiments were performed simultaneously, thus sharing the same control. Drug administration was performed on late-L4 worms unless otherwise noted. RNAi, SPD or rapamycin treatments were continued throughout life. In autophagy experiments, identical concentrations of SPD (0.2 mM) and rapamycin (50 μΜ) were administered and autophagic events were assessed at the first round of IF.
C. elegans molecular cloning and RNA interference
RNAi was performed by feeding methods 101 and was performed lifelong, starting from egg hatching. To generate the RNAi constructs, gene-specific fragments of interest were amplified by PCR directly from the C. elegans genomic DNA using appropriate primer sets. The PCR-generated fragments were initially inserted into the TOPO-pCRII vector and then sub-cloned in the pL4440 plasmid vector. The final constructs were transformed into HT115 (DE3) Escherichia coli bacteria.
For argn-1(RNAi) , a 1,409-bp fragment was amplified by using the primer set:
For_argn-1: 5′-ATGAAAAAGTCTACACAACTCGCCAGA-3′ and
Rev_argn-1: 5′-TCACATTGCTCTTGTAATTTTCTGAGATTG-3′.
For dhps-1(RNAi) , a 2,420-bp fragment was amplified by using the primer set:
For_dhps-1: 5′-ATGAGCACCAACGAAGCAGCAG-3′ and
Rev_dhps-1: 5′-ATGCTTGGCCGCCCAATACA-3′.
For odc-1(RNAi) , a 1,327-bp fragment was amplified by using the primer set:
odc-1_For: 5′-ATGATTTCTCAATTCGAAATTATTGGTGAC-3′ and
odc-1_Rev: 5′-ATCACATACATCGGCACAGGCTTC-3′.
For smd-1(RNAi) , we amplified a 1,724-bp fragment by using the primer set:
For_smd-1: 5′-ATGTCTGCCACGTCTGCCAC-3′ and
Rev_smd-1: 5′-TCCTCGTCGCTCGATGATGA-3′.
For spds-1(RNAi) , a 1,714-bp fragment was amplified by using the primer set:
For_spds-1: 5′-ATGAACAAGCTGCACAAGGGATG-3′ and
rev_spds-1: 5′-CTTGCGATGACAAAATTCCATCCTC-3′.
C. elegans mRNA quantification
Quantification of mRNA levels was performed as previously described 102 . Total mRNA was isolated from synchronized adults, lysed in 250 μl TRIzol by freeze-cracking (Invitrogen). For complementary DNA synthesis, mRNA was reverse transcribed using an iScriptTM cDNA Synthesis kit (Bio-Rad). Quantitative PCR was performed in triplicate using a Bio-Rad CFX96 Real-Time PCR system (Bio-Rad). The housekeeping gene act-3 was used as an internal normalization control. In each experiment, three technical replicates were used per sample.
For act-3 mRNA detection, the following set of primers was used:
act-3 RT Fw: 5′-ATCCGTAAGGACTTGTACGCCAAC-3′ and
act-3 RT Rev: 5′-CGATGATCTTGATCTTCATGGTTC-3′.
For smd-1 mRNA detection, the following set of primers was used:
smd-1RT_For: 5′-CTCAAACAGGATAGATGGTACCTTTACACAT-3′ and
smd-1RT_Rev: 5′-TTGTCAATACCGGCTCGCATG-3′.
For spds-1 mRNA detection, the following set of primers was used:
spds-1RT_For: 5′-GATGTGCTGGTTTTTGAGAGCACA-3′ and
spds-1RT_Rev: 5′-ATCTCCTCCACCAATGATAAGTACACGT-3′.
For dhps-1 mRNA detection, the following set of primers was used:
dhps-1RT_For: 5′-GGTGACCAGTGGGCTTCGTGAGGTAC-3′ and
dhps-1RT_Rev: 5′-TTATCATTCGGAATGAGCACATTGC-3′.
For dohh-1 mRNA detection, the following set of primers was used:
dohh-1_RTFor: 5′-ATTGCTGATCCTTCAGTCAAAGATGTCCTC-3′ and
dohh-1_RTRev: 5′-GTTGAAGCTGATGGTGTTGGATCAA-3′.
For odc-1 mRNA detection, the following set of primers was used:
odc-1RT_For: 5′-CATCTGCCTACAGGAATGCTCTTCA-3′
odc-1RT_Rev: 5′-TCACGAATTGTCTCGGCAATCTTC-3′.
For argn-1 mRNA detection, the following set of primers was used:
argn-1RT_For: 5′-AGTGCAATGTCAGGAGTTACACAGACAT-3′ and
argn-1RT_Rev: 5′-CATCAACCCAAATAAGCCCAATATCTC-3′.
For amx-3 mRNA detection, the following set of primers was used:
amx-3RT_For: 5′-TCCAGTCCGGTAGGTGAGCTT-3′ and
amx-3RT_Rev: 5′-CAGTTCAATCTAATGTTTCCCGCAGGTAT-3′.
For hpo-15 mRNA detection, the following set of primers was used:
hpo-15RT_For: 5′-ACATGATGAGAAGACTGCAAGACGCTA-3′ and
hpo-15RT_Rev: 5′-TCCACTCTTTTAGAACGTGAACACTCAGAGT-3′.
For d2023.4 mRNA detection, the following set of primers was used:
d2023.4RT_For: 5′-TGCATATTCAACATGGGTTGGGCAGTAT-3′ and
d2023.4RT_Rev: 5′-TCAACAGTGTCGTACAGAGCAATCGCAT-3′.
For iff-1 mRNA detection, the following set of primers was used:
iff-1RT_For: 5′-ATCGAAGACGGGTTCTGCTCGCT-3′ and
iff-1RT_Rev: 5′-CAGGCAGCAACGACTTGAACCAAG-3′.
C. elegans heat stress assay
Five-day-old N2 (WT) animals (after the first round of fasting) were placed at 37 °C for 6 h. After recovery overnight at 20 °C, survival was assessed. Animals that did not exhibit either touch-provoked movement or pharyngeal pumping were considered dead. Five independent experiments were performed.
C. elegans detection of autophagy
To measure autophagic flux, the C. elegans strain MAH215 (sqIs11 (lgg-1p::mCherry::GFP::lgg-1 + rol-6)) was used (LGG-1 is the C. elegans orthologue of LC3/Atg8). Adult animals after the first round of fasting were measured. We calculated the number of autolysosomes (ALs) (mCherry-positive puncta). The number of mCherry-only puncta was calculated (the total number of mCherry-positive puncta − the number of GFP-positive + mCherry-positive puncta) as previously shown 103 . Z-stack images were acquired using the LEICA TCS SP8 laser scanning confocal microscope unit. Measurements were performed in the terminal bulb and at the area where the pharyngeal lumen was best observed. Additionally, we calculated the ratio of mCherry:GFP signal by measuring mean fluorescence intensity in the head region of animals, as previously reported 104 . We also calculated SQST-1::GFP (the C. elegans orthologue of SQSTM1) positive particles using the HZ589: bpIs151 (sqst-1p::sqst-1::GFP + unc-76(+)) strain. Particles were measured in the head region of the animals. Images were acquired using the EVOS Cell Imaging Systems.
C. elegans size measurement
Five-day-old N2 (WT) animals (after the first round of fasting) were anaesthetized and observed with ×4 magnification. Images were acquired using the EVOS Cell Imaging Systems. Animal periphery was precisely circled manually using the ‘polygon selections’ tool in ImageJ and the contained area was automatically calculated.
C. elegans immunoblotting
For sample preparation, synchronous animal populations were collected after 48 h of fasting and washed in M9 buffer. After washing, two volumes of homogenization buffer (20 mM Tris, pH 7.4, 20 mM NaCl and 1 mM MgCl 2 ) containing complete mini proteinase inhibitor cocktail (Roche) in a final concentration 1× was added. One volume of beads (0.5 mm zirconium oxide beads) was added and samples were placed in a Bullet Blender Homogenizer (Model BT24M, Next Advance) for 3 min at speed 10. We used Bio-Rad protein assay (Bio-Rad, 5000006) to determine the protein concentration of the supernatant. The final sample buffer (final concentration of 62.5 mM Tris-HCl, pH 6.8, 2% SDS, 8.7% glycerol, 0.004% bromophenol blue and 120 mM DTT) was added to the supernatant. In brief, before electrophoresis, samples were boiled at 95 °C for 5 min and centrifuged at 10,000 rcf for 1 min. Protein extract corresponding to 5 µg protein was loaded onto 4–12% Bis-Tris gels and electrophoresis was performed at 90–120 V. Proteins were wet-transferred to methanol-activated 0.45-µm PVDF membranes (Roth, T830.1) at 220 mA for 60–90 min using transfer buffer (10 mM CAPS/NaOH, pH 11 and 10% methanol). After blotting, membranes were blocked with blocking solution (1% dry milk powder in TBS, pH 7.4) for 1 h and then incubated with the primary antibodies overnight at 4 °C. After three washing steps in TBST for 5 min, membranes were incubated with secondary, HRP-linked antibodies for 1 h at room temperature. After three washing steps in TBST for 5 min, signals were detected with a ChemiDoc detection system (Bio-Rad) and Clarity Western ECL Substrate (Bio-Rad) using the ‘optimal exposure’ setting. Band intensities were quantified using ImageLab v.5.2 (Bio-Rad) using the rectangular volume tool with local background adjustment. Primary antibodies were anti-hypusine (Merck, ABS1064-I, 1:1,000 dilution in blocking solution) and anti-GAPDH clone GA1R (Thermo Fisher Scientific, MA5-15738, 1:10,000 dilution in blocking solution). Secondary antibodies were HRP-linked anti-mouse IgG (Sigma, A9044, 1:10,000 dilution in blocking solution) or HRP-linked anti-rabbit IgG (Sigma, A0545, 1:10,000 dilution in blocking solution).
Fasting and caloric restriction protocols in mice
Acute fasting for polyamine measurement.
C57BL/6J:Rj male and female mice were purchased from Janvier Labs at the age of 3–4 months. Animals were housed in groups of four mice in individually ventilated cages (Tecniplast Type 2L) under specific-pathogen-free (SPF) conditions in a 14:10-h light–dark cycle with access to food (standard maintenance chow, Ssniff, cat. no. V1536) and autoclaved tap water ad libitum. Autoclaved nest material and paper houses served as cage enrichment for mice. Animals were randomly assigned to fasting or control groups. Fasting was performed overnight for 14–16 h by food withdrawal, starting between 16:00 and 17:00 (lights off at 20:00), with water ad libitum at an age of 5–6 months.
Protocol 1 (IF 16:8 )
Male C57BL/6J:Rj mice were purchased (from Janvier Labs) at the age of 16–17 months. Animals were housed in groups of four mice in individually ventilated cages under SPF conditions in a 12-h light–dark cycle with access to standard chow ad libitum until begin of the fasting training followed by intermittent fasting. Specifically, mice were subjected to 3 weeks of fasting training, thereby reducing the daily feeding time window every two or three days from 24 to 14, 12, 10 and finally 8 h, to ensure that the aged mice slowly adapted to the final intermittent fasting (IF 16:8 ) regimen, with access to food from 7:00 to 15:00 during the light phase, except at weekends, when they had access to food ad libitum. During the feeding time period, the animals had unrestricted access to food. DFMO (a kind gift from P. M. Woster, Medical University of South Carolina) aqueous solution was passed through a sterile filter and prepared fresh every week in the drinking water at 1% or 0.5% w / v . Before the experiments, the animals were randomly divided into four groups: ad libitum, ad libitum + DFMO, IF and IF + DFMO. The treatments were initiated at the age of 17–18 months. DFMO was administered at the concentration of 1% w / v for 4 weeks and then reduced to 0.5% w / v for the remainder of the experiment.
Protocol 2 (IF CR )
C57BL/6J:Rj male mice were purchased from Janvier Labs, France at the age of 18–19 months. Animals were housed in groups of four mice in individually ventilated cages (Tecniplast Type 2L) under SPF conditions in a 14:10-h light–dark cycle with access to food (standard maintenance chow, Ssniff, cat. no. V1536) and autoclaved tap water ad libitum. Autoclaved nest material and paper houses served as cage enrichment for mice. Animals were randomly assigned to fasting or control groups. Baseline calorie intake was calculated across the whole cohort over 4 weeks (4.3 g average food intake per mouse and day). Intermittent fasting was performed as described previously 50 starting at an age of ~20 month and the mice were randomly assigned to the groups. In brief, IF CR groups were fed once daily, shortly before the dark phase, with a single meal providing 70% of the cohort’s baseline calorie intake. The mice quickly consumed the allocated food portions and fasted for the remainder of the day, as manually verified in regular early morning visits. DFMO, prepared as described above, was given ad libitum at 0.25 and 0.5% ( w / v ) in the drinking water.
Protocol 3 (IF 24:24 )
Male and female mice 8–12 weeks of age were randomly assigned into four groups using randomizer.org . The control group (ad lib) was given free access to food and water during the whole experiment and received no further treatment. The feeding regimen of the IF 24:24 group involved restricting access to food for 24 h, followed by 24 h food ad libitum. The IF + DFMO group was, in addition to fasting, treated with 1% ( w / v ) DFMO, prepared as described above, in the drinking water. The SPD group was given free access to food and was treated with 3 mM SPD (Sigma, 85558; from 1 M aqueous stock, pH 7.4) in the drinking water, prepared as previously described 23 . DFMO and SPD were exchanged two and three times every week, respectively.
Protocol 4 (caloric restriction)
C57BL/6J mice were obtained from The Jackson Laboratory. Mice were bred at the National Institute on Aging (NIA) and fed chow diets (NIH-31 diet; T.9717.15, Envigo) ad libitum or were on a 30% CR, starting at 9 months of age, as described previously 31 . CR mice were fed between 6:30 and 8:30.
Transgenic mice
Male mice overexpressing human IGF1R specifically in cardiomyocytes (IGF1R tg mice) 40 and male mice expressing a dominant negative phosphoinositide 3-kinase (PI3K) p110α mutant with impaired catalytic activity restricted to cardiomyocytes (dnPI3K mice) 41 were used. In brief, a truncated p110 mutant with p85 binding domains, but lacking the kinase domain (p110Δkinase), was generated, and the p110Δkinase gene, together with FLAG epitope tag, was cloned into the αMyHC promoter construct to produce dnPI3K transgenic mice 41 . Both transgenic mouse models and their WT littermates were generated on the FVB/N genetic background and were killed at 3–9 months of age for heart tissue analysis, which included immunoblotting and polyamine measurements.
Healthspan measurements in aged mice
Various healthspan measurements were performed on IF CR mice, with adapted protocols as detailed previously 105 . The measurements were performed in the morning until noon and the operators were blinded to the group allocation. In brief, grip strength (two paws and four paws, on different days) was assessed with a grip strength meter (TSE), according to the manufacturer’s manual. The maximum grip strength of three repeats was counted and normalized to body weight of the animal on the test day. A visual frailty index was performed as previously described 106 , which assesses several phenotypical alterations occurring with age in mice. Body composition of physically restrained animals was analysed with an NMR MinspecAnalyzer (Bruker). A four-limb grid-hanging test was performed at a height of 35 cm above a cushioned cage and the time to fall was recorded manually. Body surface temperature was recorded in the lower abdomen with a handheld infra-red thermometer and the maximum of three recordings was counted.
Echocardiography in aged mice
Cardiac function and dimensions were evaluated in lightly anaesthetized mice (4–5% for induction; 1–1.5% isoflurane for maintenance) by transthoracic echocardiography using a high-resolution micro-imaging system Vevo3100 (Fujifilm VisualSonics). Animal temperature was kept at 37 °C using a temperature-controlled heating platform, on which mice were placed in a supine position with their limbs in direct contact with electrode pads for heart rate assessment. Pre-warmed ultrasound transmission gel was spread on a shaved chest to obtain cardiac tracings in the parasternal long axis using 55 MHz linear-array probe. As previously described 42 , M-mode tracings were used to evaluate cardiac walls thickness and internal left ventricular (LV) dimensions at the level of the papillary muscles during systole and diastole. The ratio between peak early filling velocity of transmitral flow (E) and the corresponding mitral valve annulus velocity (e′) was assessed using pulsed wave and tissue Doppler imaging, respectively. All measures were averaged from at least three cardiac cycles under stable conditions.
K/BxN serum transfer arthritis model
BALB/cJRj mice were purchased from Janvier Labs. Mice were kept in a temperature- and humidity-controlled facility with free access to water and a 12-h light–dark cycle. All experiments were performed using age- and sex-matched littermate controls.
After 3 weeks of intermittent fasting/treatment, 6 h after re-feeding, 100 µl serum from arthritic transgenic K/BxN mice (obtained from the animal facility of the University of Erlangen) were administered to mice via intraperitoneal injection. The severity of K/BxN serum transfer arthritis 107 was then clinically assessed by visual evaluation of swelling and redness in each paw, including the tarsus and carpus joints as previously described 108 . In short, the following scoring system was used: 0, no erythema or swelling; 1, erythema and swelling in up to two joints; 2, erythema and swelling in more than two joints or slight swelling of the ankle; 3, marked swelling of the ankle; and 4, substantial swelling of the whole paw, including toes and fingers. Scores of all paws were added together and AUCs of arthritis courses were calculated. IF and DFMO treatments were continued until the end of the experiment 24 days after serum transfer.
Detection of polyamines
Tissues were dissected and snap-frozen in liquid nitrogen after killing animals by a final blood draw (from retro-orbital plexus) under deep isoflurane narcosis followed by cervical dislocation. Serum was obtained via 30 min incubation at room temperature, followed by centrifugation at 1,000–2,000 rcf for 10 min at 4 °C. Appropriate amounts of frozen tissue (10–50 mg) were homogenized in varying volumes of 5% TCA to yield a final SPD concentration of 50–1,000 ng ml −1 (corresponds to the validated linear range of detection) in the final extract. Polyamine extraction and measurement were performed as previously described, with adapted analyte concentrations in the calibration solutions 82 (see also ‘Yeast’ section).
Immunoblotting
For protein extraction, tissue was homogenized in lysis buffer (150 mM NaCl, 20 mM Tris-HCl, pH 7.5, 1 mM EGTA, 1 mM EDTA, 1% Triton X-100, 2× Complete protease inhibitor cocktail (Roche) and 1× PhosSTOP (Roche); 400 µl lysis buffer per 30 mg of tissue) for 30 s with a Turrax (IKA T10 basic) followed by an incubation for 30 min at 4 °C with continuous rotation. The homogenized samples were centrifuged for 10 min at 20,000 rcf at 4 °C. We used a Bio-Rad protein assay (Bio-Rad 5000006) to determine the protein concentration of the supernatant. Final sample buffer (final concentration: 62.5 mM Tris-HCl, pH 6.8, 2% SDS, 8.7% glycerol, 0.004% bromophenol blue and 120 mM DTT) was added to the supernatant. In brief before electrophoresis, samples were boiled at 95 °C for 5 min and centrifuged at 10,000 rcf for 1 min. Protein extract corresponding to 20 µg protein was loaded onto 4–12% Bis-Tris gels and electrophoresis was performed at 90–120 V. Proteins were transferred to methanol-activated 0.45-μm PVDF membranes for 90 min at 220 mA using transfer buffer (10 mM CAPS pH 11 and 10% methanol). The membranes were then incubated in blocking solution (3% dry milk powder in TBS, pH 7.4) for 1 h and subsequently incubated with the primary antibody overnight at 4 °C. Membranes were washed three times with TBST for 5 min and subsequently incubated with the secondary antibody for 1 h at room temperature. After three washing steps in TBST for 5 min, signals were detected with a ChemiDoc detection system (Bio-Rad) and Clarity Western ECL Substrate (Bio-Rad) using the ‘optimal exposure’ setting. Band intensities were quantified using ImageLab v.5.2 (Bio-Rad) using the rectangular volume tool with local background adjustment. For re-probing membranes, Restore PLUS Western Blot Stripping Buffer (Thermo Fisher Scientific, 46430) was used according to the manufacturer’s protocol. Primary antibodies were anti-hypusine (Merck, ABS1064-I, 1:1,000 dilution), anti-eIF5a (BD Biosciences, 611977, 1:10,000 dilution) and anti-GAPDH clone GA1R (Thermo Fisher Scientific, MA5-15738, 1:10,000 dilution) in 1% dry milk powder in TBS, pH 7.4. Secondary antibodies were HRP-linked anti-mouse IgG (Sigma, A9044, 1:10,000 dilution in 1% dry milk powder in TBS, pH 7.4) and HRP-linked anti-rabbit IgG (Sigma, A0545, 1:10,000 dilution in 1% dry milk powder in TBS, pH 7.4).
Ethical regulations
All animal experiments were performed in accordance with national and European ethical regulation (Directive 2010/63/EU) and approved by the responsible government agencies (Bundesministerium für Wissenschaft, Forschung und Wirtschaft, BMWFW; BMWFW-66.007/0029-WF/V/3b/2017, GZ 2021-0.524.242, GZ 2022-0.137.213, BMWFW-66.010/0160-WF/V/3b/2014, BMWFW-66.010/0198-WF/V/3b/2017 and BMBWF-66.010/0042-V/3b/2018). Animal studies using protocol 3 (IF 24:24 ) and the arthritis model were approved by the local ethical committee (District Government of Lower Franconia, 55.2-2532-2-1041-15). All experiments were conducted according to the guidelines of the Federation of European Laboratory Animal Science Associations. CR mice (protocol 4) were used in accordance with protocols approved by the Institutional Animal Care and Use Committee of the NIA.
Human cells
Cell lines, media and compounds.
All culture media and supplements for human cell culture were purchased from Gibco-Life Technologies. Plasticware was purchased from Greiner Bio-One. Cells used in this study were cultured in DMEM containing 10% FBS and non-essential amino acids (Gibco, 111400) at 37 °C under 5% CO 2 . Hanks’ balanced salt solution (HBSS; Gibco,14025092) was used for starvation experiments. DFMO, spermidine, rapamycin, torin-1, aminoguanidine, bafilomycin and chloroquine were obtained from Sigma-Aldrich. The pharmacological DHS inhibitor N1-guanyl-1,7-diamine-heptane (GC7) was purchased from Merck (259545). DFMO and GC7 were prepared as sterile filtered 50 mM stocks in ddH 2 O and stored in aliquots at −20 °C.
We used human glioblastoma H4 and human osteosarcoma U2OS cells (from ATCC), either in WT form or stably expressing GFP–LC3. Generally, cells were seeded in six-well plates at approximately 1 × 10 5 cells per well. DFMO treatment was initiated after overnight attachment by exchanging the medium with freshly prepared DFMO-containing medium.
Cell line polyamine measurements
The plates were placed on ice and quickly washed with cold PBS (4 °C). Then, 500 µl of cold methanol:water (9:1 v / v , with internal standards) with 2% of sulfosalicylic acid were added to each well and the plate was stored overnight at −80 °C. Two wells at ~5 × 10 5 cells per well were scrapped and pooled for one measurement in microcentrifuge tubes. For normalization purposes, we used reference wells for cell counting.
The suspension was vortexed for 30 s and centrifuged at 15,000 rpm (21,130 rcf) for 10 min at 4 °C. After the transfer of the whole supernatant to another set of microtubes, samples were evaporated. The dried samples were spiked with 200 µl of MilliQ water before injection in UHPLC–MS. Polyamine analysis was performed by LC–MS/MS with a 1290 UHPLC (Agilent Technologies) coupled to a QQQ 6470 (Agilent Technologies) 84 .
Cell line autophagy induction and microscopy
DFMO treatment (48–72 h) was continuously performed, whereas GC7 was used acutely during starvation. Trypsinized GFP–LC3-expressing cells were seeded in 384-well black microplates (~4,000 cells per well). Starvation with HBSS or treatment with rapamycin (10 µM; 2 mM DMSO stock stored in aliquots at −20 °C) or Torin-1 (300 nM; 60 µM DMSO stock stored in aliquots at −20 °C) was initiated after overnight attachment. Before starvation experiments, the cells were gently washed with warm PBS (37 °C). DMSO at a final concentration of 0.5% was used as the control treatment for rapamycin and torin-1. Fresh DMEM containing FBS and non-essential amino acids was used as the control treatment for starvation experiments. Chloroquine (50 µM in sterile filtered ddH 2 O) was used as an autophagic flux inhibitor and added to the wells after half of the treatment time, except for Extended Data Fig. 5r,s , where bafilomycin A1 (BafA1, 100 nM) was used.
Experiments involving spermidine supplementation (Sigma, 85558; from aqueous stocks, pH 7.4) were co-treated with aminoguanidine (from aqueous stocks, 1 mM, Sigma, 396494).
After treatment, cells were fixed with 3.7% paraformaldehyde ( w / v in PBS containing 2 µg ml −1 Hoechst) for 20 min at room temperature. Microscopy images were acquired with an ImageXpress Micro Confocal microscope (Molecular Devices) equipped with a ×20 PlanApo objective (Nikon). Four view fields and six z-stacks were imaged per well. We used the Custom Module Editor functionality of the MetaXpress software (Molecular Devices) to analyse the images. First, using the nuclear Hoechst staining and cytoplasmic GFP signal, cells were segmented into nuclear and cytoplasmic regions. Then, cytoplasmic GFP–LC3 puncta were detected using automated thresholding, and their number and the cellular surface covered by GFP–LC3 puncta were analysed.
Cell line siRNA interference
U2OS cells (with or without stably expressing GFP–LC3) were seeded in 384-well imaging microplates at an approximate density of 1,000 cells per well (for autophagy measurements) or in six-well plates at an approximate density of 1 × 10 5 cells per well cells per well (for polyamine measurements). Following 1 day of attachment, cells were transfected with siRNAs against ODC1 or a cherry-picked siRNA library (Dharmacon) targeting selected genes associated with polyamine metabolism at a final concentration of 30 nM using the DharmaFECT transfection reagent (Dharmacon) according to the manufacturer’s instructions. Each gene was represented as four individual siRNAs and non-targeting siRNAs served as control (Supplementary Table 4 ). After 24 h, the medium was renewed and cells were left to adapt for an additional 24 h. Subsequently, cells were carefully washed with warm PBS and subjected to starvation with HBSS for 6 h.
Image acquisition was carried out as previously described. The obtained images were segmented and analysed with the freely available software R ( https://www.r-project.org ), integrated with the following packages: EBImage from the Bioconductor repository ( https://www.bioconductor.org ), MetaxpR ( https://github.com/kroemerlab/MetaxpR ), RBioFormats ( https://github.com/aoles/RBioFormats ), as well as MorphR ( https://github.com/kroemerlab/MorphR ). The primary region of interest was designated by a mask around the nucleus to enable cell enumeration, while a secondary cytoplasmic region of interest was employed for the segmentation of GFP–LC3 puncta. Following the exclusion of cellular debris and dead cells, the number of GFP–LC3 dots per cell was quantified and represented as fold change relative to the control.
Cell line RNA extraction and gene expression analysis
For RNA extraction, the RNeasy Plus Mini kit (cat. no. 74134, QIAGEN) was utilized. Cell pellets were stored at −80 °C before purifying RNA. Buffer RLT Plus was used to lyse the cell pellets. The lysate was centrifuged and subjected to further purification following the manufacturer’s instructions. All steps of the procedure were carried out at room temperature and all centrifugation steps were performed at 20–25 °C in a standard microcentrifuge. Approximately 1 μg total RNA was reverse transcribed using a Maxima First Strand cDNA Synthesis kit (cat. no. K1642, Thermo Fisher Scientific). Quantitative reverse transcription PCR (qRT–PCR) was conducted using PowerUp SYBR Green Master Mix (cat. no. A25776, Thermo Fisher Scientific) with a StepOnePlus Real-Time PCR System (Applied Biosystems, Thermo Fisher Scientific). The 2 −ΔΔCT method was applied for the analysis of real-time PCR data with the appropriate primers. All primers used for gene expression analysis are listed in Supplementary Table 5 .
Cell lines immunoblotting
For immunoblotting, 2–3 wells from six-well plates were washed with PBS, then collected on ice with RIPA cell lysis buffer. Cells were scraped off and incubated for 30 min at 4 °C before centrifugation for 20 min at 12,000 rpm (13,523 rcf). The protein concentration in the supernatant was determined with a Bradford assay and the samples were stored at −20 °C until further processing. In brief before electrophoresis, samples were mixed with final sample buffer (final concentration of 62.5 mM Tris-HCl, pH 6.8, 2% SDS, 8.7% glycerol, 0.004% bromophenol blue and 120 mM DTT) and boiled at 95 °C for 5 min. Then, 10 µg total protein was loaded onto 4–12% Bis-Tris gels and run at 90–120 V. Proteins were transferred to 0.45-μm PVDF membranes for 90 min at 220 mA using transfer buffer (10 mM CAPS pH 11 and 10% methanol). The membranes were then incubated in blocking solution (3% dry milk powder in TBS, pH 7.4) for 1 h and subsequently incubated with the primary antibody overnight at 4 °C. Membranes were washed three times with TBST for 5 min and subsequently incubated with the secondary antibody for 1 h at room temperature. After three washing steps in TBST for 5 min, signals were detected with a ChemiDoc detection system (Bio-Rad) and Clarity Western ECL Substrate (Bio-Rad) using the ‘optimal exposure’ setting. For re-probing membranes, Restore PLUS Western Blot Stripping Buffer (Thermo Fisher Scientific, 46430) was used according to the manufacturer’s protocol. Band intensities were quantified using ImageLab v.5.2 (Bio-Rad) using the rectangular volume tool with local background adjustment. Primary antibodies were anti-hypusine (Merck, ABS1064-I, 1:1,000 dilution), anti-GAPDH clone GA1R (Thermo Fisher Scientific, MA5-15738, 1:10,000 dilution) and anti-eIF5A (611977, BD Biosciences, 1:10,000 dilution), diluted in 1% dry milk powder in TBST. Secondary antibodies were HRP-linked anti-mouse IgG (Sigma, A9044, 1:10,000 dilution in blocking solution) or HRP-linked anti-rabbit IgG (Sigma, A0545, 1:10,000 dilution in blocking solution).
Human fasting trials
Written informed consent was obtained from all volunteers before they were enrolled in the studies, which were conducted in accordance with the principles of the Declaration of Helsinki.
The study protocol was approved by the medical council of Baden-Württemberg under the application number F-2018-118 and registered in the German Clinical Trials Register (ID DRKS00016657). The 109 participants were recruited among the in-house patients of the Buchinger Wilhelmi clinic Überlingen as previously described 109 , 110 , 111 . All participants underwent a 10 ± 3-day fasting period under medical supervision. A daily caloric intake of around 250 kcal was provided by 250 ml organic juice, 250 ml vegetable soup and 20 g honey. Serum samples were collected before the start of the fast and at the end of the 10 ± 3-day fasting period. Tubes were stored at −70 °C until further analysis.
The 1-year observational study was performed in the Buchinger Wilhelmi clinic Überlingen and described in detail previously 112 , 113 , 114 , 115 . The medical council of Baden-Württemberg and the Ethics Committee of the Charité-University Medical Center Berlin approved the study protocol under application no. EA4/054/15 that was registered in the German Clinical Trials Register (ID DRKS00010111). Blood samples before and at the end of fasting were available for a subgroup of 63 participants who fasted more than 4 days according to the Buchinger Wilhelmi fasting programme. Serum samples were stored at −70 °C.
Cohort 3 was previously described 116 , 117 . For the purpose of this study, we took a subset of study participants with available plasma samples (23 of an initial 30 performing alternate day fasting) after 12 and 36 h of fasting, stored them at −70 °C and performed targeted MS of spermidine as described below, which was previously not quantified.
Clinical data were collected during an exploratory clinical trial. Healthy participants were included by written informed consent when presenting no manifest chronic diseases nor pregnancy or lactation. Recruitment and intervention were performed on an outpatient basis at the Immanuel Hospital Berlin, where the outpatient department of the Institute of Social Medicine, Epidemiology and Health Economics of the Charité – Universitätsmedizin Berlin is seated. The study protocol was approved by the institutional review board of Charité Universitätsmedizin Berlin (ID EA1/263/20) and is registered with ClinicalTrials.gov under NCT04739852 .
Recruitment for healthy volunteers took place in August 2020. All measurements and the intervention were conducted in September 2020. Participants underwent a 5-day prolonged fasting period with a dietary energy intake of 200–350 kcal per day with vegetable broths and juices. These were followed by three days of gradual re-introduction of solid food intake, the meals containing mainly complex carbohydrates. PBMCs and blood were collected at baseline, after 1, 3 and 5 days of the fasting intervention, as well as on days 3 and 7 after re-introduction of food.
The blood of healthy volunteers was collected in EDTA-sampling tubes (Sarstedt, 02.1066.001) via venipuncture, carefully loaded on Biocoll solution (BioChrom, L6113) in Lecosep tubes (Z642843, Greiner) and centrifuged at 684 rcf for 30 min (brakeless running down). PBMCs were enriched by selecting the interphase of the Biocoll gradient. PBMCs of the interphase were washed twice with ice-cold PBS. Pellets were stored at −80 °C until they were further processed. This is the first publication of data from this cohort and the primary end points will be published elsewhere.
Detection of polyamines and other metabolites in clinical samples
Polyamine quantification from serum was performed as previously described 82 .
Cohorts 2 and 3
Serum and plasma samples were treated following a previously described protocol 84 . In brief, 50-µl samples were vortexed for 5 min with 500 µl ice-cold extraction mixture (methanol:water, 9:1, −20 °C, with a cocktail of internal standards) and then centrifuged (10 min at 15,000 rpm (21,130 rcf), 4 °C). Several fractions were then split to be analysed by LC–MS and GC–MS 84 . Polyamine analyses were performed by LC–MS/MS with a 1290 UHPLC (Agilent Technologies) coupled to a QQQ 6470 (Agilent Technologies) as previously described 85 .
All targeted treated data were merged and cleaned with a dedicated R (v.4.0) package (@Github/Kroemerlab/GRMeta).
Cohort 4 serum
For the extraction of polar metabolites from human serum samples, a mixture of methyl tert-butyl ether (Sigma-Aldrich, 650560), methanol (Biosolve, 136841) and water (Biosolve, 232141) (all LC–MS grade) was utilized in a 50:30:20 ( v : v : v ) ratio as the sample extraction buffer. The sample extraction buffer included the following internal standards: U- 13 C 15 N-labelled amino acids at a final concentration of 0.25 µM (2.5 mM in 0.1 N HCl, Cambridge Isotope Laboratories, MSK-A2-1.2), citric acid d 4 at 0.02 µg ml −1 (100 µg ml −1 in H 2 O, Sigma-Aldrich, 485438-1 G), ATP 13 C 10 at 0.1 µg ml −1 (1 mg ml −1 in 5 mM Tris-HCl, Sigma-Aldrich, 710695), AMP 13 C 10 , 15 N 5 at 0.1 µg ml −1 (1 mg ml −1 in 5 mM Tris-HCl, Sigma-Aldrich, 650676), ADP 15 N 5 at 0.1 µg ml −1 (1 mg ml −1 in 5 mM Tris-HCl, Sigma-Aldrich, 741167) and EquiSPLASH LIPIDOMIX at 0.02 µg ml −1 (100 µg ml −1 , Avanti Polar Lipids, 30731). The sample extraction buffer was freshly prepared and cooled to −20 °C before a volume of 1 ml of the chilled sample extraction buffer was added to 50 µl human serum. This mixture was incubated at 4 °C for 30 min with shaking at 1,500 rpm in a thermomixer (VWR, Thermal Shake lite). Post-incubation, the samples were centrifuged (Eppendorf, Centrifuge 5425R) at 4 °C for 10 min at 21,000 rcf and the cleared supernatant was transferred to a 2-ml tube. To this supernatant, 200 µl methyl tert-butyl ether and 150 µl water were added, followed by incubation at 15 °C for 10 min with shaking at 1,500 rpm in a thermomixer. A subsequent centrifugation at 15 °C for 10 min at 16,000 rcf facilitated phase separation. Approximately 650 µl of the upper lipid-containing phase was transferred to a 1.5-ml tube (not included in this study). The remaining polar metabolite extract, approximately 600 µl, was left in the 2-ml tube after removal of the residual lipid phase. The polar metabolite extract was then distributed in two 1.5-ml tubes and dried in a SpeedVac concentrator (LaboGene, Scan Speed 40) at 20 °C at 1,000 rpm. The resulting dried metabolite pellets were stored at −80 °C until further analysis.
Cohort 4 PBMCs
For the extraction of polar metabolites from human PBMCs, a mixture of acetonitrile (Biosolve, 012041), methanol and water (all LC–MS-grade) was utilized in a 40:40:20 ( v : v : v ) ratio as the sample extraction buffer. The sample extraction buffer included the following internal standards: U- 13 C 15 N-labelled amino acids at a final concentration of 0.25 µM (2.5 mM in 0.1 N HCl, Cambridge Isotope Laboratories, MSK-A2-1.2), citric acid d 4 at 0.02 µg ml −1 (100 µg ml −1 in H 2 O, Sigma-Aldrich, 485438-1 G), ATP 13 C 10 at 0.1 µg ml −1 (1 mg ml −1 in 5 mM Tris-HCl, Sigma-Aldrich, 710695), AMP 13 C 10 , 15 N 5 at 0.1 µg ml −1 (1 mg ml −1 in 5 mM Tris-HCl, Sigma-Aldrich, 650676), and ADP 15 N 5 at 0.1 µg ml −1 (1 mg ml −1 in 5 mM Tris-HCl, Sigma-Aldrich, 741167). The sample extraction buffer was freshly prepared and cooled to −20 °C before a volume of 1 ml of the chilled sample extraction buffer was added to the individual PBMC pellets. To dissolve the pellets, samples were subjected to 3–5 pulses of sonication using a tip sonicator (Bandelin, Sonopuls). This mixture was then incubated at 4 °C for 30 min with shaking at 1,500 rpm in a thermomixer. Post-incubation, the samples were centrifuged at 4 °C for 10 min at 21,000 rcf and the cleared supernatant was transferred to a 1.5-ml tube and the pellet (protein pellet) was set aside. The polar metabolite extract was then dried in a SpeedVac concentrator at 20 °C at 1,000 rpm. The resulting dried metabolite pellets were stored at −80 °C until further analysis. The residual liquid was removed from the protein pellet and the pellet was left to air dry at room temperature for protein quantification using the Pierce 660 nm BCA protein quantification kit (Thermo Fisher Scientific, 22660) according to the manufacturer’s instructions. For protein extraction, pellets were re-suspended in a protein extraction buffer (50 mM Tris-HCl, pH 8.5, 1% SDS and 140 mM NaCl), homogenized with metal balls (3.5 mm) using a mixer mill (Retsch, MM 400) and diluted in an appropriate volume of protein extraction buffer before assay measurement.
Cohort 4: semi-targeted LC–high-resolution MS-based analysis of amine-containing metabolites
The LC–high-resolution MS analysis of amine-containing compounds was performed using a QE-Plus high-resolution mass spectrometer coupled to a Vanquish UHPLC chromatography system (Thermo Fisher Scientific). In brief, dried sample extracts were dissolved in 150 µl LC–MS-grade water for 10 min at 4 °C in a shaker at 1,500 rpm. After centrifugation, 50 µl of the extracts were mixed with 25 µl of 100 mM sodium carbonate (Sigma-Aldrich), followed by the addition of 25 µl 2% ( v / v ) benzoylchloride (Sigma-Aldrich) in acetonitrile (ULC/MS grade, Biosolve). The derivatized samples were thoroughly mixed and kept at a temperature of 20 °C until analysis.
For the LC–high-resolution MS analysis, 1 µl of the derivatized sample was injected onto a 100 × 2.1-mm HSS T3 UPLC column (Waters). The flow rate was set to 400 µl min −1 using a binary buffer system consisting of buffer A (10 mM ammonium formate (Sigma-Aldrich) and 0.15% ( v / v ) formic acid (Sigma-Aldrich) in ULC/MS-grade water (Biosolve)). Buffer B consisted of acetonitrile (ULC/MS-grade, Biosolve). The column temperature was set to 40 °C, while the LC gradient was: 0% B at 0 min, 0–15% B 0–4.1 min; 15–17% B 4.1–4.5 min; 17–55% B 4.5–11 min; 55–70% B 11–11.5 min, 70–100% B 11.5–13 min; B 100% 13–14 min; 100–0% B 14–14.1 min; 0% B 14.1–19 min; 0% B. The mass spectrometer (Q-Exactive Plus) was operating in positive ionization mode recording the mass range m/z 100–1,000. The heated electrospray ionisation (ESI) source settings of the mass spectrometer were spray voltage 3.5 kV, capillary temperature 300 °C, sheath gas flow 60 a.u., aux gas flow 20 a.u. at 330 °C and the sweep gas was set to 2 a.u. The RF lens was set to a value of 60.
LC–MS data analysis was performed using the open-source software El Maven 118 (v.0.12.0). For this purpose, Thermo raw mass spectra files were converted to mzML format using MSConvert 119 (v.3.0.22060, Proteowizard). The identity of each compound was validated by authentic reference compounds, which were measured at the beginning and the end of the sequence. For data analysis, the area of the protonated (M + nBz + H) + (where, nBz indicates the number of benzoyl moieties attached to each compound) mass peaks of every required compound were extracted and integrated using a mass accuracy of <5 ppm and a retention time tolerance of <0.05 min compared with the independently measured reference compounds. Peak areas were then normalized to internal standards and to the protein content (PBMCs) or to the median of the total ion count (serum). For cohort 4, data were further normalized across the average values of the time points per participant.
Protein extraction and capillary immunoblotting (cohort 4)
Protein extracts from PBMCs were obtained by lysing cells (approx. 1–2 × 10 6 cells) in 80 µl T-PER extraction buffer (Thermo Scientific, 78510), supplemented with protease (Sigma, P2714) and phosphatase (Roche, 04906837001) inhibitor cocktail. Protein extracts (1–2 µg) were separated by capillary electrophoresis on Jess (ProteinSimple) using 12–230 kDa cartridges (ProteinSimple, SM-W004) and the following primary antibodies for detection: anti-hypusine (1:50 dilution, ABS1064-I, Merck), anti-eIF5a (1:50 dilution, 611977, BD Biosciences) and anti-ODC1 (1:50 dilution, PA5-21362, Thermo Scientific). The signals were normalized to total protein using the protein normalization module (DM-PN02, ProteinSimple).
Statistics and reproducibility
All data were checked for normality using QQ plots and histograms. In the case of nonparametric data distribution and feasibility, data were log 2 -transformed and tested again for normality. In the case of persistent nonparametric data distribution, appropriate statistical nonparametric tests were used. A post hoc Holm-Šídák’s test (false discovery rate 5%) or Dunn’s test were used to account for multiple testing within a dataset. For statistical analysis of the survival curves, we used log-rank tests with post hoc Bonferroni corrections for multiple testing. Outlier analysis using the ROUT method (Q = 0.1–0.5%) and outlier exclusion were performed on the metabolite measurements of human biological samples. We used GraphPad Prism 10.1.0 ( www.graphpad.com ) for general data visualization and statistics. All applied statistical tests are listed in the figure legends. For specific details of the OMICS data analyses see the respective Methods sub-sections. Sample size corresponds to either individual measurements of animals or human participants, or groups of cells (yeast and human cell lines) or flies. All data points shown are independent samples and all samples on immunoblots were biologically independent. Immunoblots used for quantification are in the source data. All experimental replications yielded comparable outcomes.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The MS yeast proteomics raw data together with the processing log files have been deposited to the ProteomeXchange Consortium via the PRIDE ( http://www.ebi.ac.uk/pride ) partner repository with the dataset identifier PXD035909 . The metabolomics and all other data are included in Source Data. All other data supporting the findings of this study are available from the corresponding author on reasonable request. Source data are provided with this paper.
Green, C. L., Lamming, D. W. & Fontana, L. Molecular mechanisms of dietary restriction promoting health and longevity. Nat. Rev. Mol. Cell Biol . https://doi.org/10.1038/s41580-021-00411-4 (2021).
Fontana, L. The scientific basis of caloric restriction leading to longer life. Curr. Opin. Gastroenterol. 25 , 144–150 (2009).
Article CAS PubMed Google Scholar
Francesco, A. D., Germanio, C. D., Bernier, M. & de. Cabo, R. A time to fast. Science 362 , 770–775 (2018).
Article PubMed PubMed Central Google Scholar
Hofer, S. J., Carmona-Gutierrez, D., Mueller, M. I. & Madeo, F. The ups and downs of caloric restriction and fasting: from molecular effects to clinical application. EMBO Mol. Med . https://doi.org/10.15252/emmm.202114418 (2021).
Mitchell, S. J. et al. Daily fasting improves health and survival in male mice independent of diet composition and calories. Cell Metab. 29 , 221–228.e3 (2019).
Trepanowski, J. F. et al. Effect of alternate-day fasting on weight loss, weight maintenance, and cardioprotection among metabolically healthy obese adults: a randomized clinical trial. JAMA Intern. Med. 177 , 930–938 (2017).
de Cabo, R. & Mattson, M. P. Effects of intermittent fasting on health, aging, and disease. N. Engl. J. Med. 381 , 2541–2551 (2019).
Article PubMed Google Scholar
Longo, V. D. & Panda, S. Fasting, circadian rhythms, and time-restricted feeding in healthy lifespan. Cell Metab. 23 , 1048–1059 (2016).
Article CAS PubMed PubMed Central Google Scholar
Longo, V. D., Di Tano, M., Mattson, M. P. & Guidi, N. Intermittent and periodic fasting, longevity and disease. Nat. Aging 1 , 47–59 (2021).
Wilhelmi de Toledo, F., Grundler, F., Bergouignan, A., Drinda, S. & Michalsen, A. Safety, health improvement and well-being during a 4 to 21-day fasting period in an observational study including 1422 subjects. PLoS ONE 14 , e0209353 (2019).
Most, J., Tosti, V., Redman, L. M. & Fontana, L. Calorie restriction in humans: an update. Ageing Res. Rev. 39 , 36–45 (2017).
Madeo, F., Zimmermann, A., Maiuri, M. C. & Kroemer, G. Essential role for autophagy in life span extension. J. Clin. Invest. 125 , 85–93 (2015).
Aman, Y. et al. Autophagy in healthy aging and disease. Nat. Aging 1 , 634–650 (2021).
Deretic, V. & Kroemer, G. Autophagy in metabolism and quality control: opposing, complementary or interlinked functions? Autophagy 18 , 283–292 (2022).
Partridge, L., Fuentealba, M. & Kennedy, B. K. The quest to slow ageing through drug discovery. Nat. Rev. Drug Discov. 19 , 513–532 (2020).
Klionsky, D. J. et al. Autophagy in major human diseases. EMBO J. 40 , e108863 (2021).
Levine, B. & Kroemer, G. Biological functions of autophagy genes: a disease perspective. Cell 176 , 11–42 (2019).
Collier, J. J. et al. Developmental consequences of defective ATG7-mediated autophagy in humans. N. Engl. J. Med. 384 , 2406–2417 (2021).
Bjedov, I. et al. Fine-tuning autophagy maximises lifespan and is associated with changes in mitochondrial gene expression in Drosophila . PLoS Genet. 16 , e1009083 (2020).
Fernández, Á. F. et al. Disruption of the beclin 1-BCL2 autophagy regulatory complex promotes longevity in mice. Nature 558 , 136–140 (2018).
Pyo, J.-O. et al. Overexpression of Atg5 in mice activates autophagy and extends lifespan. Nat. Commun. 4 , 2300 (2013).
Kaeberlein, M. Spermidine surprise for a long life. Nat. Cell Biol. 11 , 1277–1278 (2009).
Eisenberg, T. et al. Induction of autophagy by spermidine promotes longevity. Nat. Cell Biol. 11 , 1305–1314 (2009).
Eisenberg, T. et al. Cardioprotection and lifespan extension by the natural polyamine spermidine. Nat. Med. 22 , 1428–1438 (2016).
Zhang, H. et al. Polyamines control eIF5A hypusination, TFEB translation, and autophagy to reverse B cell senescence. Mol. Cell 76 , 110–125.e9 (2019).
Hofer, S. J. et al. Mechanisms of spermidine-induced autophagy and geroprotection. Nat. Aging https://doi.org/10.1038/s43587-022-00322-9 (2022).
Alsaleh, G. et al. Autophagy in T cells from aged donors is maintained by spermidine and correlates with function and vaccine responses. eLife 9 , e57950 (2020).
Kiechl, S. et al. Higher spermidine intake is linked to lower mortality: a prospective population-based study. Am. J. Clin. Nutr. 108 , 371–380 (2018).
Beck, T. & Hall, M. N. The TOR signalling pathway controls nuclear localization of nutrient-regulated transcription factors. Nature 402 , 689–692 (1999).
Wullschleger, S., Loewith, R. & Hall, M. N. TOR signaling in growth and metabolism. Cell 124 , 471–484 (2006).
Pomatto, L. C. D. et al. Deletion of Nrf2 shortens lifespan in C57BL6/J male mice but does not alter the health and survival benefits of caloric restriction. Free Radic. Biol. Med . 152 , 650–658 (2020).
Onodera, J. & Ohsumi, Y. Autophagy is required for maintenance of amino acid levels and protein synthesis under nitrogen starvation*. J. Biol. Chem. 280 , 31582–31586 (2005).
May, A. I., Prescott, M. & Ohsumi, Y. Autophagy facilitates adaptation of budding yeast to respiratory growth by recycling serine for one-carbon metabolism. Nat. Commun. 11 , 5052 (2020).
Liu, K., Sutter, B. M. & Tu, B. P. Autophagy sustains glutamate and aspartate synthesis in Saccharomyces cerevisiae during nitrogen starvation. Nat. Commun. 12 , 57 (2021).
Saxton, R. A., Chantranupong, L., Knockenhauer, K. E., Schwartz, T. U. & Sabatini, D. M. Mechanism of arginine sensing by CASTOR1 upstream of mTORC1. Nature 536 , 229–233 (2016).
Suzuki, S. W., Onodera, J. & Ohsumi, Y. Starvation induced cell death in autophagy-defective yeast mutants is caused by mitochondria dysfunction. PLoS ONE 6 , e17412 (2011).
Bertram, P. G. et al. Tripartite regulation of Gln3p by TOR, Ure2p, and phosphatases*. J. Biol. Chem. 275 , 35727–35733 (2000).
Hardwick, J. S., Kuruvilla, F. G., Tong, J. K., Shamji, A. F. & Schreiber, S. L. Rapamycin-modulated transcription defines the subset of nutrient-sensitive signaling pathways directly controlled by the Tor proteins. Proc. Natl Acad. Sci. USA 96 , 14866–14870 (1999).
Schmelzle, T. & Hall, M. N. TOR, a central controller of cell growth. Cell 103 , 253–262 (2000).
McMullen, J. R. et al. The insulin-like growth factor 1 receptor induces physiological heart growth via the phosphoinositide 3-kinase(p110α) pathway. J. Biol. Chem. 279 , 4782–4793 (2004).
Shioi, T. et al. The conserved phosphoinositide 3-kinase pathway determines heart size in mice. EMBO J. 19 , 2537–2548 (2000).
Abdellatif, M. et al. Fine-tuning cardiac insulin-like growth factor 1 receptor signaling to promote health and longevity. Circulation 145 , 1853–1866 (2022).
Noda, T. & Klionsky, D. J. The quantitative Pho8Delta60 assay of nonspecific autophagy. Methods Enzymol. 451 , 33–42 (2008).
Longo, V. D., Shadel, G. S., Kaeberlein, M. & Kennedy, B. Replicative and chronological aging in Saccharomyces cerevisiae . Cell Metab. 16 , 18–31 (2012).
Tsukada, M. & Ohsumi, Y. Isolation and characterization of autophagy-defective mutants of Saccharomyces cerevisiae . FEBS Lett. 333 , 169–174 (1993).
Kaeberlein, M. et al. Regulation of yeast replicative life span by TOR and Sch9 in response to nutrients. Science 310 , 1193–1196 (2005).
Powers, R. W., Kaeberlein, M., Caldwell, S. D., Kennedy, B. K. & Fields, S. Extension of chronological life span in yeast by decreased TOR pathway signaling. Genes Dev. 20 , 174–184 (2006).
Yuan, H., Wu, S.-X., Zhou, Y.-F. & Peng, F. Spermidine inhibits joints inflammation and macrophage activation in mice with collagen-induced arthritis. J. Inflamm. Res 14 , 2713–2721 (2021).
Abdellatif, M., Rainer, P. P., Sedej, S. & Kroemer, G. Hallmarks of cardiovascular ageing. Nat. Rev. Cardiol. 20 , 754–777 (2023).
Acosta-Rodríguez, V. et al. Circadian alignment of early onset caloric restriction promotes longevity in male C57BL/6J mice. Science 376 , 1192–1202 (2022).
Whitehead, J. C. et al. A clinical frailty index in aging mice: comparisons with frailty index data in humans. J. Gerontol. Ser. A 69 , 621–632 (2014).
Article Google Scholar
Park, M. H. & Wolff, E. C. Hypusine, a polyamine-derived amino acid critical for eukaryotic translation. J. Biol. Chem. 293 , 18710–18718 (2018).
Liang, Y. et al. eIF5A hypusination, boosted by dietary spermidine, protects from premature brain aging and mitochondrial dysfunction. Cell Rep. 35 , 108941 (2021).
Lubas, M. et al. eIF5A is required for autophagy by mediating ATG3 translation. EMBO Rep. 19 , e46072 (2018).
Dever, T. E. & Ivanov, I. P. Roles of polyamines in translation. J. Biol. Chem. 293 , 18719–18729 (2018).
Ivanov, I. P. et al. Polyamine control of translation elongation regulates start site selection on the antizyme inhibitor mRNA via ribosome queuing. Mol. Cell 70 , 254–264.e6 (2018).
Kaushik, S. et al. Autophagy and the hallmarks of aging. Ageing Res. Rev. 72 , 101468 (2021).
Frankel, L. B. EIF5A mediates autophagy via translation of ATG3. Autophagy 14 , 1288–1289 (2018).
Liao, C.-Y. et al. The autophagy inducer spermidine protects against metabolic dysfunction during overnutrition. J. Gerontology: Ser. A https://doi.org/10.1093/gerona/glab145 (2021).
Tain, L. S. et al. Longevity in response to lowered insulin signaling requires glycine N -methyltransferase-dependent spermidine production. Aging Cell 19 , e13043 (2020).
Obata, F. & Miura, M. Enhancing S -adenosyl-methionine catabolism extends Drosophila lifespan. Nat. Commun. 6 , 8332 (2015).
Gao, A. W. et al. Identification of key pathways and metabolic fingerprints of longevity in C. elegans . Exp. Gerontol. 113 , 128–140 (2018).
Kimura, K. D., Tissenbaum, H. A., Liu, Y. & Ruvkun, G. daf-2, an insulin receptor-like gene that regulates longevity and diapause in Caenorhabditis elegans . Science 277 , 942–946 (1997).
Kenyon, C., Chang, J., Gensch, E., Rudner, A. & Tabtiang, R. A C. elegans mutant that lives twice as long as wild type. Nature 366 , 461–464 (1993).
Ulgherait, M. et al. Circadian autophagy drives iTRF-mediated longevity. Nature 598 , 353–358 (2021).
Zwighaft, Z. et al. Circadian clock control by polyamine levels through a mechanism that declines with age. Cell Metab. 22 , 874–885 (2015).
Pietrocola, F. et al. Caloric restriction mimetics enhance anticancer immunosurveillance. Cancer Cell 30 , 147–160 (2016).
Al-Habsi, M. et al. Spermidine activates mitochondrial trifunctional protein and improves antitumor immunity in mice. Science 378 , eabj3510 (2022).
Holbert, C. E., Cullen, M. T., Casero, R. A. & Stewart, T. M. Polyamines in cancer: integrating organismal metabolism and antitumour immunity. Nat. Rev. Cancer 22 , 467–480 (2022).
Janke, C. et al. A versatile toolbox for PCR-based tagging of yeast genes: new fluorescent proteins, more markers and promoter substitution cassettes. Yeast 21 , 947–962 (2004).
Noda, T., Matsuura, A., Wada, Y. & Ohsumi, Y. Novel system for monitoring autophagy in the yeast Saccharomyces cerevisiae . Biochem. Biophys. Res. Commun. 210 , 126–132 (1995).
Eisenberg, T. et al. Nucleocytosolic depletion of the energy metabolite acetyl-coenzyme a stimulates autophagy and prolongs lifespan. Cell Metab. 19 , 431–444 (2014).
Kainz, K. et al. Methods to assess autophagy and chronological aging in yeast. Methods Enzymol. 588 , 367–394 (2017).
Schindelin, J. et al. Fiji: an open-source platform for biological-image analysis. Nat. Methods 9 , 676–682 (2012).
Carmona-Gutierrez, D. et al. The flavonoid 4,4′-dimethoxychalcone promotes autophagy-dependent longevity across species. Nat. Commun. 10 , 651 (2019).
Burtner, C. R., Murakami, C. J., Kennedy, B. K. & Kaeberlein, M. A molecular mechanism of chronological aging in yeast. Cell Cycle 8 , 1256–1270 (2009).
Steffen, K. K., Kennedy, B. K. & Kaeberlein, M. Measuring replicative life span in the budding yeast. J. Vis. Exp. https://doi.org/10.3791/1209 (2009).
Kaeberlein, M., Kirkland, K. T., Fields, S. & Kennedy, B. K. Sir2-independent life span extension by calorie restriction in yeast. PLoS Biol. 2 , e296 (2004).
Péli-Gulli, M.-P., Sardu, A., Panchaud, N., Raucci, S. & De Virgilio, C. Amino acids stimulate TORC1 through Lst4-Lst7, a GTPase-activating protein complex for the rag family GTPase Gtr2. Cell Rep. 13 , 1–7 (2015).
Zhang, F. et al. Tissue-specific landscape of metabolic dysregulation during ageing. Biomolecules 11 , 235 (2021).
Pang, Z. et al. MetaboAnalyst 5.0: narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. https://doi.org/10.1093/nar/gkab382 (2021).
Magnes, C. et al. Polyamines in biological samples: rapid and robust quantification by solid-phase extraction online-coupled to liquid chromatography–tandem mass spectrometry. J. Chromatogr. A 1331 , 44–51 (2014).
Costa-Machado, L. F. et al. Peripheral modulation of antidepressant targets MAO-B and GABAAR by harmol induces mitohormesis and delays aging in preclinical models. Nat. Commun. 14 , 2779 (2023).
Grajeda-Iglesias, C. et al. Oral administration of Akkermansia muciniphila elevates systemic antiaging and anticancer metabolites. Aging (Albany NY) 13 , 6375–6405 (2021).
Durand, S. et al. in Methods in Cell Biology Vol. 164 (eds Kepp, O. & Galluzzi, L.) Ch. 11 (Academic Press, 2021).
Meier, F. et al. diaPASEF: parallel accumulation–serial fragmentation combined with data-independent acquisition. Nat. Methods 17 , 1229–1236 (2020).
Demichev, V., Messner, C. B., Vernardis, S. I., Lilley, K. S. & Ralser, M. DIA-NN: neural networks and interference correction enable deep proteome coverage in high throughput. Nat. Methods 17 , 41–44 (2020).
Kamburov, A., Stelzl, U., Lehrach, H. & Herwig, R. The ConsensusPathDB interaction database: 2013 update. Nucleic Acids Res. 41 , D793–D800 (2013).
Meldal, B. H. M. et al. Analysing the yeast complexome—the Complex Portal rising to the challenge. Nucleic Acids Res. 49 , 3156–3167 (2021).
R Core Team. R: a language and environment for statistical computing. https://www.R-project.org (R Foundation for Statistical Computing, 2023).
Venables, W. N. & Ripley, B. D. Modern Applied Statistics with S (Springer, 2002).
Mardia, K. V., Kent, J. T. & Bibby, J. M. Multivariate Analysis (Academic Press, 1979).
Wickham, H. Ggplot2 (Springer International Publishing, 2016).
Kolde, R. Pheatmap: pretty heatmaps. R package version 1 , 726 (2012).
Google Scholar
Schroeder, S. et al. Dietary spermidine improves cognitive function. Cell Rep. 35 , 108985 (2021).
Ja, W. W. et al. Prandiology of Drosophila and the CAFE assay. Proc. Natl Acad. Sci. USA 104 , 8253–8256 (2007).
Salvi, D., Waggoner, J., Temlyakov, A. & Wang, S. A graph-based algorithm for multi-target tracking with occlusion. In 2013 IEEE Workshop on Applications of Computer Vision (WACV) 489–496 (2013).
Brenner, S. The genetics of Caenorhabditis elegans . Genetics 77 , 71–94 (1974).
Honjoh, S., Yamamoto, T., Uno, M. & Nishida, E. Signalling through RHEB-1 mediates intermittent fasting-induced longevity in C. elegans . Nature 457 , 726–730 (2009).
Robida-Stubbs, S. et al. TOR signaling and rapamycin influence longevity by regulating SKN-1/Nrf and DAF-16/FoxO. Cell Metab. 15 , 713–724 (2012).
Kamath, R. S., Martinez-Campos, M., Zipperlen, P., Fraser, A. G. & Ahringer, J. Effectiveness of specific RNA-mediated interference through ingested double-stranded RNA in Caenorhabditis elegans . Genome Biol. 2 , RESEARCH0002 (2001).
CAS PubMed Google Scholar
Lionaki, E. et al. Mitochondrial protein import determines lifespan through metabolic reprogramming and de novo serine biosynthesis. Nat. Commun. 13 , 651 (2022).
Chang, J. T., Hansen, M. & Kumsta, C. in Aging: Methods and Protocols (ed. Curran, S. P.) 187–200 (Springer, 2020).
Tezil, T. et al. Lifespan-increasing drug nordihydroguaiaretic acid inhibits p300 and activates autophagy. NPJ Aging Mech. Dis. 5 , 1–11 (2019).
Bellantuono, I. et al. A toolbox for the longitudinal assessment of healthspan in ageing mice. Nat. Protoc. 15 , 540 (2020).
Kane, A. E., Keller, K. M., Heinze-Milne, S., Grandy, S. A. & Howlett, S. E. A murine frailty index based on clinical and laboratory measurements: links between frailty and pro-inflammatory cytokines differ in a sex-specific manner. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 74 , 275 (2019).
Article CAS Google Scholar
Christensen, A. D., Haase, C., Cook, A. D. & Hamilton, J. A. K/BxN serum-transfer arthritis as a model for human inflammatory arthritis. Front. Immunol. 7 , 213 (2016).
Kienhöfer, D. et al. No evidence of pathogenic involvement of cathelicidins in patient cohorts and mouse models of lupus and arthritis. PLoS ONE 9 , e115474 (2014).
Ravanidis, S. et al. Fasting-mediated metabolic and toxicity reprogramming impacts circulating microRNA levels in humans. Food Chem. Toxicol. 152 , 112187 (2021).
Grundler, F. et al. Interplay between oxidative damage, the redox status, and metabolic biomarkers during long-term fasting. Food Chem. Toxicol. 145 , 111701 (2020).
Wilhelmi de Toledo, F. et al. Influence of long-term fasting on blood redox status in humans. Antioxid. 9 , 496 (2020).
Grundler, F. et al. Long-term fasting improves lipoprotein-associated atherogenic risk in humans. Eur. J. Nutr. 60 , 4031–4044 (2021).
Grundler, F., Mesnage, R., Michalsen, A. & Wilhelmi de Toledo, F. Blood pressure changes in 1610 subjects with and without antihypertensive medication during long-term fasting. J. Am. Heart. Assoc. https://doi.org/10.1161/JAHA.120.018649 (2020).
Toledo, F. W., de, Grundler, F., Bergouignan, A., Drinda, S. & Michalsen, A. Safety, health improvement and well-being during a 4 to 21-day fasting period in an observational study including 1422 subjects. PLoS ONE 14 , e0209353 (2019).
Drinda, S. et al. Effects of periodic fasting on fatty liver Index-A prospective observational study. Nutrients 11 , E2601 (2019).
Stekovic, S. et al. Alternate day fasting improves physiological and molecular markers of aging in healthy, non-obese humans. Cell Metab. 30 , 462–476.e5 (2019).
Tripolt, N. J. et al. Intermittent fasting (alternate day fasting) in healthy, non-obese adults: protocol for a cohort trial with an embedded randomized controlled pilot trial. Adv. Ther. 35 , 1265–1283 (2018).
Agrawal, S. et al. in High-Throughput Metabolomics: Methods and Protocols (ed. D’Alessandro, A.) 301–321 (Springer, 2019).
Chambers, M. C. et al. A cross-platform toolkit for mass spectrometry and proteomics. Nat. Biotechnol. 30 , 918–920 (2012).
Download references
Acknowledgements
We thank all participants of the clinical trials and the medical staff for helping conduct the trials. We are grateful to P. M. Woster (Medical University of South Carolina, USA) for providing DFMO used in the fly and mouse experiments. Further, we are grateful for the technical assistance provided by M. Hausl (Joanneum Research HEALTH, Graz, Austria), L. Opriessnig (Institute of Molecular Biosciences (IMB), University of Graz, Austria), S. Fasching (Institute of Pharmaceutical Sciences, University of Graz, Austria), and all members of the animal facility teams at IMB and Biomedical Research, as well as K. Zierler, R. Schreiber (IMB, University of Graz, Austria), S. Kurat and M. Prassl (Biomedical Research, Medical University of Graz, Austria) for organizational help and assistance with the animal licences. Moreover, we thank A. Sauvat (Centre de Recherche des Cordeliers, France) for helping with cellular segmentation analysis and A. Gross (Gregor Mendel Institute of Molecular Biology, Vienna, Austria) for comments on the manuscript. This research was funded in part by the Austrian Science Fund (FWF) (grants DOC-50, F3012, W1226, P29203, P29262, P27893 and P31727 (F.M.); grants P27637-B28 and I3301-MINOTAUR (S.S.); grants P28854, I3792, DOC-130 and DK-MCD W1226 (T.M.); grants P33957 and TAI6021000 (T.E.)). For the purpose of open access, the author has applied a CC BY public copyright licence to any Author Accepted Manuscript version arising from this submission. Further funding was provided by Agence National de la Recherche (ANR) – Projets blancs (G.K.); Austrian Federal Ministry of Education, Science and Research and the University of Graz grants ‘Unkonventionelle Forschung-InterFast’, ‘Fast4Health’, ‘flysleep’ (BMWFW-80.109/0001-WF/V/3b/2015) (F.M.); Austrian Research Promotion Agency (F.F.G.) grants 864690 and 870454 (T.M.); BioTechMed-Graz (Flagship project DYNIMO) (U.S., S.M. and T.M.); BioTechMed-Graz (Flagship project EPIAge) (F.M.); BioTechMed-Graz (Flagship project INTERACD + ) (S.S. and T.E.). Cancer Research ASPIRE Award from the Mark Foundation (G.K.); Cancéropôle Ile-de-France (G.K.); Deutsche Forschungsgemeinschaft grant SI 849/14-1 (Project-ID 445178831) (S.J.S. and Y.L.); Elior (G.K.). Equipex Onco-Pheno-Screen 21-ESRE-0028 (G.K.); European Joint Programme on Rare Diseases (G.K.); European Research Council grant ‘ERC-GA695190-MANNA’ (N.T.); European Union Horizon 2020 FETOPEN, project Dynamic-GA‐863203 (N.T.); European Union Research Excellence Hub changing-GA‐101087071 (N.T.); European Union Horizon 2020 Project Oncobiome (G.K.); Fondation ARC (O.K.); German Research Council, DFG 672032 (N.C.G.); German Research Foundation, DFG, FOR2886/2-TP08 and EXC2167-390884018 (M.H.H.); Greek National Research Infrastructure project BIOIMAGING‐GR-MIS5002755 (N.T.); Hellenic Foundation for Research and Innovation and the General Secretariat for Research and Technology grant 1898 (N.T.); IdEx Université de Paris ANR-18-IDEX-0001 (G.K.); Longevity Impetus Grant (M.A.M.); Institut National du Cancer (G.K. and O.K.); Institut Universitaire de France (G.K.); Integrative Metabolism Research Center Graz, the Austrian Infrastructure Program 2016/2017 (T.M.); LabEx Immuno-Oncology ANR-18-IDEX-0001 (G.K.); Ligue Contre le Cancer, Équipe Labellisée (G.K.); Medical University of Graz (Flagship project VASC-HEALTH) (S.S. and M.A.); NIH 5P20GM121176 (M.A.M.); NIH R01AG07077601 (M.A.M.). In part supported by the Intramural Research Program at the NIA, National Institutes of Health (R.d.C); RHU Immunolife (G.K.); Seerave Foundation (G.K.); SIRIC Cancer Research and Personalized Medicine, CARPEM (G.K.); SIRIC Stratified Oncology Cell DNA Repair and Tumor Immune Elimination SOCRATE (G.K.); Styrian Government (Zukunftsfonds, doc.fund program) (T.M.); Swiss National Science Foundation 310030_166474/184671 (C.D.V.); Swiss National Science Foundation 310030_184781 (J.D.) and The French National Cancer Institute INCA PLBIO21-286 (O.K.). The authors acknowledge the financial support by the University of Graz and the University of Graz Field of Excellence BioHealth (U.S., T.E. and F.M.)
Author information
These authors contributed equally: Nektarios Tavernarakis, Guido Kroemer, Frank Madeo.
Authors and Affiliations
Institute of Molecular Biosciences, NAWI Graz, University of Graz, Graz, Austria
Sebastian J. Hofer, Martina Bergmann, Andreas Zimmermann, Melanie I. Mueller, Alexander Nartey, Mara Waltenstorfer, Sarah Enzenhofer, Isabella Faimann, Verena Gschiel, Lukas Pein, Jelena Tadic, Andrea Jerkovic, Tobias Eisenberg & Frank Madeo
Field of Excellence BioHealth, University of Graz, Graz, Austria
Sebastian J. Hofer, Andreas Zimmermann, Jelena Tadic, Ulrich Stelzl, Tobias Eisenberg & Frank Madeo
BioTechMed Graz, Graz, Austria
Sebastian J. Hofer, Andreas Zimmermann, Mahmoud Abdellatif, Sarah Masser, Jelena Tadic, Thomas R. Pieber, Simon Sedej, Tobias Madl, Ulrich Stelzl, Tobias Eisenberg & Frank Madeo
Centre de Recherche des Cordeliers, Équipe Labellisée par la Ligue Contre le Cancer, Université de Paris Cité, Sorbonne Université, Inserm U1138, Institut Universitaire de France, Paris, France
Sebastian J. Hofer, Mahmoud Abdellatif, Sylvère Durand, Giulia Cerrato, Hui Pan, Fanny Aprahamian, Nitharsshini Nirmalathasan, Oliver Kepp & Guido Kroemer
Metabolomics and Cell Biology Platforms, Gustave Roussy Cancer Center, Université Paris Saclay, Villejuif, France
Institute of Molecular Biology and Biotechnology, Foundation for Research and Technology – Hellas, Heraklion, Greece
Ioanna Daskalaki, Ilias Gkikas & Nektarios Tavernarakis
Department of Biology, School of Sciences and Engineering, University of Crete, Heraklion, Greece
Ioanna Daskalaki & Ilias Gkikas
Department of Dermatology, Allergy and Venerology, University of Lübeck, Lübeck, Germany
Jasna Friščić & Markus H. Hoffmann
Institute for Systemic Inflammation Research, University of Lübeck, Lübeck, Germany
Division of Cardiology, Medical University of Graz, Graz, Austria
Mahmoud Abdellatif & Simon Sedej
Department of Biology, University of Fribourg, Fribourg, Switzerland
Raffaele Nicastro, Michael Stumpe, Joern Dengjel & Claudio De Virgilio
Institute of Pharmaceutical Sciences, Pharmaceutical Chemistry, University of Graz, Graz, Austria
Sarah Masser, Elisabeth Annerer & Ulrich Stelzl
Neurohomeostasis Research Group, Department of Psychiatry and Psychotherapy, University Hospital Bonn, Bonn, Germany
Thomas Bajaj, Christine Niemeyer, Daniel E. Heinz & Nils C. Gassen
Institute for Biology and Genetics, Freie Universität Berlin, Berlin, Germany
YongTian Liang & Stephan J. Sigrist
Cluster of Excellence, NeuroCure, Berlin, Germany
Department of Biochemistry and Molecular Biology, University of New Mexico Health Sciences Center, Albuquerque, NM, USA
Christine E. Robbins & Mark A. McCormick
Research Unit Integrative Structural Biology, Otto Loewi Research Center, Medicinal Chemistry, Medical University of Graz, Graz, Austria
Hansjörg Habisch & Tobias Madl
Max Planck Institute for Biology of Ageing, Cologne, Germany
Frederik Dethloff
Buchinger Wilhelmi Clinic, Überlingen, Germany
Franziska Grundler & Françoise Wilhelmi de Toledo
Institute of Social Medicine, Epidemiology and Health Economics, corporate member of Freie Universität Berlin and Humboldt-Universität, Charité-Universitätsmedizin, Berlin, Germany
Daniela A. Koppold, Anika Rajput Khokhar & Andreas Michalsen
Department of Pediatrics, Division of Oncology and Hematology, Charité – Universitätsmedizin Berlin, Corporate Member of Freie Universität Berlin and Humboldt-Universität zu Berlin, Berlin, Germany
Daniela A. Koppold
Department of Internal Medicine and Nature-based Therapies, Immanuel Hospital Berlin, Berlin, Germany
Daniela A. Koppold & Andreas Michalsen
Department of Dermatology, Venereology and Allergology, Charité – Universitätsmedizin Berlin, corporate member of Freie Universität Berlin and Humboldt-Universität zu Berlin, Berlin, Germany
Anika Rajput Khokhar
Interdisciplinary Metabolic Medicine Trials Unit, Division of Endocrinology and Diabetology, Medical University of Graz, Graz, Austria
Norbert J. Tripolt, Harald Sourij & Thomas R. Pieber
Division of Endocrinology and Diabetology, Department of Internal Medicine, Medical University of Graz, Graz, Austria
HEALTH - Institute for Biomedical Research and Technologies, Joanneum Research Forschungsgesellschaft, Graz, Austria
Thomas R. Pieber & Christoph Magnes
Experimental Gerontology Section, Translational Gerontology Branch, National Institute on Aging, National Institutes of Health, Baltimore, MD, USA
Rafael de Cabo
Institute of Physiology, Faculty of Medicine, University of Maribor, Maribor, Slovenia
Simon Sedej
Division of Basic Sciences, School of Medicine, University of Crete, Heraklion, Greece
Nektarios Tavernarakis
Institut du Cancer Paris CARPEM, Department of Biology, Hôpital Européen Georges Pompidou, AP-HP, Paris, France
Guido Kroemer
You can also search for this author in PubMed Google Scholar
Contributions
Conceptualization was by S.J.H. and F.M. Methodology was conducted by S.J.H., I.D., J.F., A.Z., M.A., R.N., S.M., S.D., Y.L., F.D., C.N. and H.H. Formal analysis was carried out by S.J.H., I.D., J.F., A.Z., M.A., R.N., S.M., S.D., Y.L., F.D., C.N., H.H. and T.M. Investigation was by S.J.H., I.D., M.B., J.F., A.Z., M.I.M., M.A., R.N., S.M., S.D., A.N., M.W., S.E., I.F., V.G., T.B., C.N., I.G., L.P., G.C., H.P., Y.L., J.T., A.J., F.A., C.E.R., N.N., H.H., E.A., F.D., M.S., D.E.H. and M.A.M. Resources were by F.G., F.W.T., D.A.K., A.R.K., A.M., N.J.T., H.S., T.R.P., R.d.C., C.M., O.K., J.D., S.J.S., N.C.G., S.S., T.M., C.D.G., U.S., M.H.H., T.E., N.T., G.K. and F.M. Data curation was carried out by S.J.H., I.D., J.F., A.Z., M.A., R.N., S.M., S.D., Y.L., H.H. and T.M. Writing of the original draft was by G.K., S.J.H. and F.M. Writing, review and editing was carried out by all authors. Visualization was conducted by S.J.H., I.D., A.Z., M.A., S.M. and Y.L. Supervision was by S.J.H., M.A.M., O.K., J.D., S.J.S., N.C.G., S.S., T.M., C.D.V., U.S., M.H.H., T.E., N.T., G.K. and F.M. Project administration was carried out by S.J.H. and F.M. Funding acquisition was by M.A., R.d.C., M.A.M., J.D., S.J.S., N.C.G., S.S., T.M., C.D.V., U.S., M.H.H., T.E., N.T., G.K. and F.M.
Corresponding authors
Correspondence to Nektarios Tavernarakis , Guido Kroemer or Frank Madeo .
Ethics declarations
Competing interests.
O.K., G.K. and F.M. are cofounders of Samsara Therapeutics, a company that develops novel pharmacological autophagy inducers. F.M. and T.E. have equity interests in and are advisors of The Longevity Labs. G.K. is a scientific co-founder of everImmune, Osasuna Therapeutics and Therafast Bio. G.K. has been holding research contracts with Daiichi Sankyo, Eleor, Kaleido, Lytix Pharma, PharmaMar, Osasuna Therapeutics, Samsara Therapeutics, Sanofi, Tollys and Vascage. G.K. is in the scientific advisory boards of Hevolution, Institut Servier and Rejuveron Life Sciences. F.G. and F.W.T. are employees of the Buchinger Wilhelmi Development und Holding, Überlingen. D.K., A.R.K. and A.M. are members of the steering committee of the Medical Association for Fasting and Nutrition (ÄGHE). D.K. and A.M. have also co-founded the Academy of Integrative Fasting, an institution for the qualification of medical staff in clinical fasting applications. D.K. serves as a consultant for a mobile application on intermittent fasting (FASTIC) as well as a company producing plant-based supplements (FENOU). A.M. is also a co-founder of the SALUFAST company and serves as a consultant for Lanserhof. All other authors declare no competing interests.
Peer review
Peer review information.
Accepted without review at Nature Cell Biology .
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended data fig. 1 changes in polyamine levels in starving organisms..
(a) Polyamine levels of WT BY4741 yeast shifted to water for the indicated times. Data normalized to the mean of the control group at every time point. N = 5 biologically independent samples (yeast cultures). (b) Polyamine levels of glucose-restricted WT BY4741 after the indicated times. Data normalized to the mean of the control group at every time point. N = 5 biologically independent samples (yeast cultures). (c) Polyamine levels of young male w 1118 flies fasted for 12 or 24 hours (starting at 20:00). Data normalized to the ad libitum group at every time point. Refed = 12 hours refeeding after 24 hours of fasting. N = 7 biologically independent samples (groups of flies). (d) Changes in body weight of young female and male w 1118 flies fasted or kept ad lib for the indicated times, starting at 20:00. Data normalized to each ad lib group. P values against each sex’s ad lib group. Refed = 12 hours refeeding after 24 hours of fasting. N = 5(Refed female fasted), 6(24 h female ad lib; Refed female ad lib; Refed male ad lib), 7(12 h, 24 h female fasted; 12 h, 24 h male; Refed male fasted) biologically independent samples (groups of flies). (e) Changes in body weight of young male C57BL/6 mice fasted or kept ad lib for 14-16 hours overnight. N = 6 mice. (f-i) Relative polyamine levels of young male C57BL/6 mice fasted or kept ad lib for 14-16 hours overnight. BAT = brown adipose tissue, WAT = white adipose tissue. FC = fold change to means of ad lib N = 3(ORN Hippocampus; PUT Muscle fasted), 4(PUT WAT fasted), 5(rest) mice. (j) Changes in body weight of young female C57BL/6 mice fasted or kept ad lib for 14-16 hours overnight. N = 8 mice. (k-n) Relative polyamine levels of young female C57BL/6 mice fasted or kept ad lib for 14-16 hours overnight. BAT = brown adipose tissue, WAT = white adipose tissue. FC = fold change to means of ad lib. N = 7(ORN Heart ad lib), 8(rest) mice. (o-q) Relative polyamine levels of male and female C57BL/6 mice kept ad lib or calorie-restricted (30 %), starting at 9 months of age, until the age of 17 or 21 months. (o) N = 4(Fasted Spleen), 6(Fasted Kidney, Serum, Heart; SPM Fasted Liver), 7(Ad lib Spleen; Fasted Muscle, Liver rest), 8(Ad lib Kidney, Liver), 10(Ad lib Serum), 11(Ad lib Heart, Muscle) mice. (p) N = 3(CR Serum SPM), 4(CR Liver, Serum rest), 5(CR Kidney, Spleen), 6(Ad lib Serum; CR Heart, Muscle), 7(), 8(Ad lib Liver, Muscle), 9(Ad lib Kidney, Heart, Spleen) mice. (q) N = 3(Ad lib Liver), 4(Ad lib Kidney, Spleen), 6(Ad lib Serum, Heart, Muscle; CR Kidney, Spleen), 7(CR Heart, Muscle), 8(CR Liver, Serum) mice. (r-s) Polyamine levels in HBSS-starved human U2OS and H4 cells. N = 4 biologically independent samples. (t) Relative mRNA expression levels of polyamine-relevant genes in 6 hours starved U2OS cells. N = 3(ARG1, OAZ1, ODC1, SAT1, ATP13A2, ATP13A3, MYC, YAP, TAZ), 4(SRM, SMS, PAOX, MAT2A, AMD1, GNMT) biologically independent samples. (u) Serum SPD levels after fasting in cohort 1, stratified by sex, depicted as mean with S.E.M and violin plots, showing median and quartiles as lines. N = 67(female), 38(male). (v-x) Serum SPD levels after fasting in cohort 1 do not correlate with age, pre-fasting baseline body mass index (BMI) or body weight loss (body weight ratio). N = 99(Body weight ratio), 102(BMI), 105(Age) volunteers. (y) Relative SPD levels in human plasma from cohort 3 during IF. N = 22(12 h), 23(36 h) volunteers. (z) ODC1 protein levels in human PBMCs after increasing fasting times, measured by capillary electrophoresis. Refeeding = day 3 after re-introduction of food. N = 15(5 d), 16(3d RF), 17(rest) volunteers. Statistics : [A-D,R-S,U,Z] Two-way ANOVA with Holm-Šídák’s multiple comparisons test. [E,J,T] Two-tailed Student’s t -tests. [F-I, K-Q] Two-way ANOVA with FDR correction (Two-stage step-up method by Benjamini, Krieger and Yekutieli, Q = 0.05). [Y] Two-tailed Student’s t -test. [V-X] Simple linear regression analysis. Bar and line graphs show the mean ± s.e.m. Heatmaps show means. * P < 0.05, ** P < 0.01, *** P < 0.001, # P < 0.2. Source numerical data are available in source data.
Extended Data Fig. 2 Spermidine supplementation corrects metabolome disturbances in ∆ spe1 yeast cells.
(a, b) Heatmap (group means) and PCA of S. cerevisiae WT and ∆ spe1 metabolomes after 6 hours -N, with or without 100 µM SPD. Unassigned NMR signals are labelled according to their NMR chemical shift. N = 6 biologically independent samples (yeast cultures). (c) Volcano plot showing significantly different metabolites in ∆ spe1 after 6 hours -N, with or without 100 µM SPD. Two-tailed Student’s t -tests with FDR-corrected P values < 0.05, FC (fold change) >1.2. N = 6 biologically independent samples (yeast cultures). (d) Metabolite set enrichment analysis based on KEGG pathways of significantly different metabolites from [C] (raw P -values < 0.2). (e) Selected metabolites from [A], focusing on amino acid metabolism and the TCA cycle. N = 6 biologically independent samples (yeast cultures). Statistics: [E] Two-way ANOVA with Holm-Šídák’s multiple comparisons test. Bar graphs show the mean ± S.E.M. Asterisks indicate raw P -values. *<0.05, **<0.01, ***<0.001. Source numerical data are available in source data.
Extended Data Fig. 3 Spermidine is required for the correct shutdown of TORC1 and autophagy regulation in N-starving yeast.
(a) Proteome change in S. cerevisiae WT BY4741 and Δ spe 1 strains under specified condition treatments following a 6-hour culture in control or -N media with or without 100 µM SPD supplementation. Differential expression (Z-score) of proteins involved in the TORC complex, from the proteome analysis. N = 6 biologically independent samples (yeast cultures). (b) Polyamine levels of WT BY472 treated with rapamycin (40 nM) for the indicated times. Data normalized to the mean of the DMSO control group at every time point. N = 5 biologically independent samples (yeast cultures). (c) Polyamine and precursor levels in cardiac tissue of young, male control, transgenic IGF1 tg or dnPI3K mice. N = 5(WT), 8(IGF1R tg ), 11(dnPI3K) mice. (d) Yeast WT and Δ spe 1 strains under specified condition treatments following a 6-hour culture in control or -N media with or without 100 µM SPD. Differential expression (Z-Score) of proteins involved in autophagy, from the proteome analysis in Supplementary Fig. 1c . N = 6 biologically independent samples (yeast cultures). (e) Differential regulation of autophagy-relevant proteins in ∆ spe1 cells. The averaged log2-transformed fold change (FC) of individual proteins (-N compared to control medium) was calculated for all conditions. The graph depicts the differences of these log2(FC) between ∆ spe1 and WT cells (red triangles), as well as ∆ spe1 treated with 100 µM SPD and WT cells (blue dots). nd=not detected. N = 6 biologically independent samples (yeast cultures). (f) Z-scores of the top 20 dysregulated proteins, as identified in [E]. N = 6 biologically independent samples (yeast cultures). Statistics : [A,B,F] Two-way ANOVA with Holm-Šídák’s multiple comparisons test. [C] Two-way ANOVA with FDR correction (Two-stage step-up method by Benjamini, Krieger and Yekutieli, Q = 0.05). Heatmaps show means. Bar and line graphs show the mean ± s.e.m. * P < 0.05, ** P < 0.01, *** P < 0.001, # P < 0.2. Source numerical data are available in source data.
Extended Data Fig. 4 SPE1, the yeast ODC1 homologue, is required for starvation- and rapamycin-induced autophagy.
(a) Representative immunoblots of yeast WT and ∆s pe1 GFP-Atg8 after 6 hours -N, assessed for GFP and GAPDH. (b) Quantifications of [B]. N = 7(∆ spe1 -N), 8 (rest) biologically independent samples (yeast cultures). (c) Representative immunoblots of yeast WT and ∆s pe1 GFP-Atg8 after 24 hours -N, assessed for GFP and GAPDH. (d) Quantifications of [C]. N = 7(∆ spe1 -N), 8(rest) biologically independent samples (yeast cultures). (e) Representative images of the categories used for categorization of GFP-Atg8 signals as shown and quantified in [F-G]. DIC = differential interference contrast, PI = propidium iodide staining for dead cells. (f) Representative images of yeast WT and ∆ spe1 GFP-Atg8 cells 6 hours after -N. Scale bar = 5 µm. (g) Blinded manual quantification of the autophagic status in microscopy images of WT and ∆ spe1 GFP-Atg8 cells 6 hours after -N. N = 6 biologically independent samples (yeast cultures). (h) ALP activity (RFU/µg) from Pho8∆N60 assay normalized to each CTL group after 24 hours -N. N = 7(WT), 6(∆ spe1 ) biologically independent samples (yeast cultures). (i) Representative immunoblots of yeast WT and ∆s pe1 GFP-Atg8 after 6 hours rapamycin treatment (40 nM), assessed for GFP and GAPDH. (j) Quantifications of [I]. N = 12 biologically independent samples (yeast cultures). (k) Representative images of yeast WT and ∆ spe1 GFP-Atg8 cells 6 hours after rapamycin (40 nM) treatment. Scale bar = 5 µm. (l) Blinded manual quantification of the autophagic status in microscopy images of WT and ∆ spe1 GFP-Atg8 cells 6 hours after rapamycin (40 nM) treatment. N = 6 biologically independent samples (yeast cultures). (m) Representative immunoblots of yeast WT BY4742 and ∆s pe1 GFP-Atg8 after 6 hours rapamycin treatment (40 nM), with and without 100 µM SPD, assessed for GFP and GAPDH. (n) Quantifications of [M]. N = 6 biologically independent samples (yeast cultures). Statistics : Two-way ANOVA with Holm-Šídák’s multiple comparisons test. Bar graphs show the mean ± s.e.m. Source numerical data and unprocessed blots are available in source data.
Extended Data Fig. 5 ODC1 is required for starvation-induced autophagy in H4 and U2OS cells.
(a) DFMO treatment (48 hours, 100 µM) affects the polyamine profile of H4 and U2OS cells in vitro . Polyamine levels normalized to cell line-specific control (0 µM DFMO). N = 5(H4 0, 100 µM DFMO), 4(rest) biologically independent samples. (b) SPD levels in DFMO-treated (48 hours, 100 µM) U2OS and H4 cells after 6 hours starvation. N = 6 biologically independent samples. (c) Quantification of surface area covered by GFP-LC3 dots in 6 hours starved U2OS GFP-LC3 cells treated with or without 100 µM DFMO for 3 days, as depicted in Fig. 3m , normalized to the control condition. N = 6 biologically independent experiments. (d) Representative images of human H4 GFP-LC3 cells starved for 3 hours in HBSS (with or without chloroquine [CQ] for 1.5 hours before fixation) after three days of 100 µM DFMO treatment. Scale bar = 10 µm. (e, f) Quantification of cytosolic GFP-LC3 dots and surface area covered by GFP-LC3 dots from [D], normalized to the control condition. N = 6 biologically independent experiments. (g) SPD levels in U2OS cells treated with DFMO (100 µM) and SPD (10 µM) after 6 hours of starvation. Aminoguanidine (1 mM) was added to all conditions. N = 6 biologically independent samples. (h-j) Representative images and quantifications of human U2OS GFP-LC3 cells starved for 6 hours in HBSS (with or without chloroquine [CQ] for 3 hours before fixation) after three days of 100 µM DFMO treatment in combination with or without 10 µM SPD. Aminoguanidine (1 mM) was added to all conditions. Scale bar = 10 µm. N = 12 biologically independent experiments. (k-m) Representative images and quantifications of human H4 GFP-LC3 cells starved for 3 hours in HBSS (with or without CQ for 1.5 hours before fixation) after three days of 100 µM DFMO treatment in combination with or without 10 µM SPD. Aminoguanidine (1 mM) was added to all conditions. Bar = 10 µm. N = 12 biologically independent experiments. (n) Polyamine levels are depleted 48 hours after ODC1 knockdown via siRNAs in U2OS cells. N = 4 biologically independent samples. (o-q) Representative images and quantifications of human U2OS GFP-LC3 cells starved for 6 hours in HBSS (with or without CQ for 3 hours before fixation) after three days of ODC1 knockdown. Scale bar = 10 µm. N = 8 biologically independent experiments. (r, s) Representative images and quantifications of human U2OS GFP-LC3 cells starved for 6 hours in HBSS or treated with Rapamycin (10 µM) or Torin-1 (300 nM). SPD was added in ascending concentrations to test for synergistic effects with starvation. AG = aminoguanidine (1 mM). N = 4(Starv.+AG + 1.25, 2.5, 320, 640, 1280 µM SPD), 8(Starv.+AG; Starv+AG + 5-160 µM SPD), 16(Rapa, Torin-1, BafA1), 32(CTL) biologically independent samples. Statistics : [A-C,E-G,I-J,L-N,P-Q] Two-way ANOVA with Holm-Šídák’s multiple comparisons test. [S] One-way ANOVA with Holm-Šídák’s multiple comparisons test Heatmaps show means. Bar graphs show the mean ± s.e.m. Source numerical data are available in source data.
Extended Data Fig. 6 Fasting-induced autophagy in worms requires odc-1.
(a) Relative mRNA expression levels of polyamine-relevant genes in 24 hours fasted C. elegans . N = 4( hpo-15 ), 5( argn-1 , smd-1 , amx-3, d2023.4 ), 6( odc-1 , spds-1 ) biologically independent experiments. (b) Knockdown efficiency of odc-1 mRNA by feeding bacteria expressing odc-1(RNAi) for 3 days in C. elegans . N = 3 biologically independent experiments. (c) Representative fluorescence images of the head region of young C. elegans MAH215 (sqIs11 [lgg-1p::mCherry::GFP::lgg-1 + rol-6]) fasted for two days and fed control or odc-1(RNAi) expressing bacteria. Autophagic activity is indicated by a shift to the red spectrum due to fluorescence quenching of the pH-sensitive-GFP by the acidic environment of the autolysosome. Scale bar = 50 μm. (d) Quantification of the ratio of the mean fluorescence intensity of mCherry/GFP signals, as depicted in [C]. Note that the experiment and statistics were performed together with the dhps-1(RNAi) groups in Extended Data Fig. 10e . N = 121(CTL ad lib), 127(CTL fasted), 113( Odc-1(RNAi) ad lib), 125( Odc-1(RNAi) fasted) worms. (e) Representative fluorescence images of the head region of young C. elegans SQST-1::GFP fasted for two days and fed control or odc-1(RNAi) . Autophagic activity is indicated by a decrease in the number of GFP-positive particles. Scale bar = 50 μm. (f) Quantification of the SQST-1::GFP particles in the head region, as depicted in [E]. Note that the experiment and statistics were performed together with the dhps-1(RNAi) groups in Extended Data Fig. 10g . N = 133(CTL ad lib), 86(CTL fasted), 109( odc-1(RNAi) ad lib), 86( odc-1(RNAi) fasted) worms. (g) Representative images of the head region of young C. elegans MAH215 (sqIs11 [lgg-1p::mCherry::GFP::lgg-1 + rol-6]) fasted for two days and fed control or odc-1(RNAi) with and without 0.2 mM SPD. Autolysosomes (ALs) appear as mCherry-positive puncta. Scale bar = 50 μm. (h) Quantification of ALs as depicted in [G]. N = 63(CTL), 42(CTL fasted), 52(CTL + SPD), 54( odc-1 (RNAi)), 48( odc-1 (RNAi) fasted), 50( odc-1 (RNAi)+SPD, odc-1 (RNAi)+SPD fasted) worms. (i) Representative images of the head region of young C. elegans MAH215 (sqIs11 [lgg-1p::mCherry::GFP::lgg-1 + rol-6]) with or without 50 µM rapamycin and fed control or odc-1(RNAi) . Autolysosomes (ALs) appear as mCherry-positive puncta. Scale bar = 50 μm. (j) Quantification of ALs as depicted in [I]. N = 63(CTL), 62(CTL+Rapa), 54( odc-1 (RNAi)), 53( odc-1 (RNAi)+Rapa) worms. Statistics : [A] Mann-Whitney with Holm-Šídák’s multiple comparisons test. [B] Two-tailed Student’s t -test. [D,F,H,J] Kruskal-Wallis test with Dunn’s multiple comparison test. Heatmaps show means. Bar graphs show the mean ± s.e.m. * P < 0.05, # P < 0.2. Source numerical data are available in source data.
Extended Data Fig. 7 Polyamine synthesis is important for IF-induced benefits in flies and worms.
(a) Daily and nightly food consumption of D. melanogaster w 1118 after 10, 30 and 50 days of IF 12:12 . N = 11(Day 30 IF Day), 12(Day 30 Ad lib), 16(Day 50 Ad lib Night), 18(Day 10), 20(Day 50 IF Day), 22(Day 50 Ad lib Day) biologically independent samples (groups of flies). (b) DFMO feeding affects the polyamine profile of female w 1118 flies during ad lib and 24 hours fasting. N = 6(Ad lib 10 µM DFMO; Fasted 0, 10 µM DFMO), 7(Ad lib 0, 0.1, 1 µM DFMO; Fasted 0.1, 1 µM DFMO) biologically independent samples (groups of flies). (c) Lifespan of male w 1118 flies fed standard food with or without 10 mM DFMO and subjected to IF. N = 210(Ad lib), 206(IF), 199(Ad lib+DFMO), 212(IF + DFMO) flies. (d) Food consumption of 10-day old female and male w 1118 flies, fed control or food containing 10 mM DFMO, during the first 7 cycles of IF. N = 8(0 mM Day IF), 9(rest) biologically independent samples (groups of 5 flies per N). (e) DFMO feeding does not influence body weight changes after 24 hours fasting. N = 7 biologically independent samples (groups of flies). (f) Lifespan of heterozygous Odc1 MI10996 /+ flies subjected to IF. The IF + SPD group received 5 mM SPD via agar during the night. N = 119( Odc1 MI10996 /+ ad lib), 117( Odc1 MI10996 /+ IF), 113( Odc1 MI10996 /+ IF + SPD) worms. (g) Knockdown of odc-1 does not affect worm size under ad lib or IF 48:48 conditions, compared to CTL RNAi. Note that the experiment and statistics were performed together with the dhps-1(RNAi) groups in Extended Data Fig. 10h . N = 56(CTL ad lib), 66(CTL IF), 63( Odc-1(RNAi) ad lib), 78( Odc-1(RNAi) IF) worms. (h) Knockdown efficiency of respective mRNAs by feeding bacteria expressing argn-1 , spds-1 , or smd-1 RNAi in C. elegans . N = 2( argn-1 , smd-1 ), 3( spds-1 ) biologically independent experiments. (i) Lifespan of C. elegans N2 fed control (CTL) or RNAi against odc-1 , with or without continuous 0.2 mM SPD feeding during IF. Note that the experiments and statistics were performed together with the groups depicted in [J]. N = 429(CTL ad lib), 523(CTL IF), 437(CTL ad lib+SPD),466(IF + SPD), 330( odc-1(RNAi) ad lib), 575( odc-1(RNAi) IF), 497( odc-1(RNAi) ad lib+SPD), 605( odc-1(RNAi ) IF + SPD) worms. (j) Lifespan under 50 µM rapamycin treatment of C. elegans N2 fed control (CTL) or RNAi against odc-1 . Note that the experiments and statistics were performed together with the groups depicted in [I]. N = 429(CTL), 423(Rapa), 330( odc-1(RNAi) , 330( odc-1(RNAi) Rapa) worms. Statistics : [A,D] Two-tailed Student’s t -test (per time point). [B,E,G] Two-way ANOVA with Holm-Šídák’s multiple comparisons test. [F,I,J] Log-rank test with Bonferroni correction. [H] Two-tailed Student’s t -test. Heatmaps show means. Bar graphs show the mean ± s.e.m. Source numerical data are available in source data.
Extended Data Fig. 8 Cardiac profiling by echocardiography of aging mice during IF, with and without DFMO.
(a) Representative echocardiography-derived mitral pulsed wave and tissue Doppler tracings in aged mice treated as outlined in Fig. 5a . (b) Representative echocardiography derived left ventricular (LV) M-mode tracings. (c) LV remodelling index. N = 13(Ad lib), 15(IF), 9(Ad lib + DFMO), 8(IF + DFMO) mice. (d) LV ejection fraction (LVEF). N = 12(Ad lib), 15(IF), 9(Ad lib + DFMO), 8(IF + DFMO) mice. (e) Heart rate. N = 13(Ad lib), 15(IF), 9(Ad lib + DFMO), 8(IF + DFMO) mice. (f) Body weight at the time of cardiac profiling. N = 13(Ad lib), 15(IF), 9(Ad lib + DFMO), 8(IF + DFMO) mice. (g) Body weight throughout the intervention. N = 15(Ad lib; Ad lib + DFMO),16(IF; IF + DFMO IF) mice (at week 0). (h) Food intake per mouse throughout the intervention. N = 3(Week 4 IF + DFMO), 4(rest) cages. (i) Grip strength (all limbs) normalized to body weight. N = 18(CTL ad lib), 15(CTL IF), 13(0.25% DFMO), 12(0.5% DFMO ad lib), 16(0.5% DFMO IF) mice. (j) Latency to fall in a 4-limb grid hanging test. N = 18(CTL ad lib), 11(CTL IF), 13(0.25% DFMO ad lib), 12(0.25% DFMO IF), 12(0.5% DFMO ad lib), 16(0.5% DFMO IF) mice. (k) Body weight. N = 18(CTL ad lib), 15(CTL IF), 14(0.25% DFMO ad lib), 13(0.25% DFMO IF), 12(0.5% DFMO ad lib), 16(0.5% DFMO IF) mice. (l) Fat-to-lean mass ratio. N = 18(CTL ad lib), 15(CTL IF), 13(0.25% DFMO), 12(0.5% DFMO ad lib), 16(0.5% DFMO IF) mice. (m) Abdominal surface temperature. N = 18(CTL ad lib), 15(CTL IF), 14(0.25% DFMO ad lib), 13(0.25% DFMO IF), 12(0.5% DFMO ad lib), 16(0.5% DFMO IF) mice. (n) Sex-stratified arthritis scoring of data presented in Fig. 5h-i upon injection of serum from K/BxN mice in young male and female BALB/cJRj mice treated as outlined in Fig. 5g . N = 4 mice. (o) Genes previously connected to the cellular effects of SPD were knocked out via siRNAs in U2OS GFP-LC3 cells and tested for starvation-induced autophagy. Selected representative images after 6 hours of starvation. (p) 48 hours after siRNA knockdown, U2OS GFP-LC3 cells were starved for 6 hours. The graph depicts the normalized individual FC to control conditions of GFP-LC3 dots for every gene knockdown or control condition (means ± S.E.M.). FC 1 represents the starvation-induced increase in GFP-LC3 dots in the control condition. Highlighted genes were significantly different from the control. N = 17(CTL), (Dharmafect control), 9(rest) biologically independent samples. Statistics : [C-N] Two-way ANOVA with Holm-Šídák’s multiple comparisons test. [P] One-way ANOVA with Holm-Šídák’s multiple comparisons test. Line graph shows the mean ± s.e.m. Bar and line graphs show the mean ± S.E.M. Source numerical data are available in source data.
Extended Data Fig. 9 Fasting increases hypusination of eIF5A.
(a) Hypusine levels are increased after 24 hours -N in yeast BY4741 GFP-Atg8 cells, but not in ∆ spe1 . Representative immunoblot. (b) Quantification of [A]. N = 7(∆ spe1 -N), 8(rest) biologically independent samples (yeast cultures). (c) Representative immunoblot of WT and ∆ spe1 , with and without 100 µM SPD after 6 hours -N, assessed for hypusine and GAPDH. (d) Quantification of [C]. N = 6 biologically independent samples (yeast cultures). (e) SPD supplementation does not further increase hypusination in BY4741 GFP-Atg8 yeast. Representative immunoblot. (f) Quantification of [E]. N = 4 biologically independent samples (yeast cultures). (g) Representative maximum projection images of confocal microscopy images of male w 1118 fly central brain regions probed for eIF5A and hypusine by immunofluorescence. Prior to dissection, the flies were fasted for 0 (ad lib) or 12 hours, starting at 8 pm. Scale bar = 50 µm. (h) Quantification of signal intensities as shown in [G]. N = 30 fly brains. (i) Representative immunoblot of female Drosophila w 1118 head extracts, assessed for hypusine and actin after 12 and 24 hours overnight fasting. RF = 12 hours refeeding. (j) Quantification of [I]. N = 6 biologically independent samples (fly head lysates). (k) Representative immunostaining microscopy images against hypusine after 12 hours fasting of female WT w 1118 and Odc1 MI10996 /+ fly brains. Scale bar = 50 µm. (l) Quantification of [K]. N = 11(ad lib), 12(fasted) fly brains. (m) Immunoblots of cardiac tissue from young male transgenic IGF1R tg or dnPI3K mice and their age-matched WT controls. Heart lysates were assessed for hypusine, total eIF5A and GAPDH. (n) Quantification of [M]. N = 6(WT), 7(IGF1R tg ), 3(dnPI3K) mice. (o) Representative immunoblot of human U2OS cells starved for 6 hours and assessed for hypusine, total eIF5A and GAPDH. (p) Quantification of [O]. N = 6 biologically independent samples. (q) Hypusine levels after five days of fasting in isolated human PBMCs of cohort 4, stratified by sex. N = 12(female baseline), 11(female fasted), 5 (male baseline), 4(male fasted) volunteers. (r-t) eIF5A H levels after five days of fasting in isolated human PBMCs of cohort 4 do not correlate with age, pre-fasting baseline BMI or body weight loss (body weight ratio). N = 15 volunteers. Statistics : [B,D,L,NP,Q] Two-way ANOVA with Holm-Šídák’s multiple comparisons test. [F] One-way ANOVA with Holm-Šídák’s multiple comparisons test. [H,J] Two-tailed Student’s t -test with Holm-Šídák’s multiple comparisons test. [R-T] Simple linear regression. Bar graphs show the mean ± s.e.m. Source numerical data and unprocessed blots are available in source data.
Extended Data Fig. 10 Hypusination is important for fasting-induced autophagy in worms and human cells.
(a) Feeding of dhps-1(RNAi) expressing bacteria reduces dhps-1 mRNA expression in C. elegans N2 worms. N = 2 biologically independent experiments. (b) Representative confocal images of the head region of young C. elegans MAH215 (sqIs11 [lgg-1p::mCherry::GFP::lgg-1 + rol-6]) fasted for two days and fed control or dhps-1(RNAi) expressing bacteria. Autolysosomes (ALs) appear as mCherry-positive puncta. Scale bar = 50 μm. (c) Quantification of ALs as depicted in [B]. Note that the experiment and statistics were performed together with the odc-1(RNAi) groups in Fig. 3p . N = 11(CTL ad lib), 26(CTL fasted), 10( dhps-1(RNAi) ad lib), 21( dhps-1(RNAi) fasted) worms. (d) Representative fluorescence images of the head region of young C. elegans MAH215 (sqIs11 [lgg-1p::mCherry::GFP::lgg-1 + rol-6]) (LGG-1 is the C. elegans ortholog of LGG-1/Atg8) fasted for two days and fed control or dhps-1(RNAi) expressing bacteria. Autophagic activity is indicated by a shift to the red spectrum due to fluorescence quenching of the pH-sensitive-GFP by the acidic environment of the autolysosome. Scale bar = 50 μm. (e) Quantification of the ratio of the mean fluorescence intensity of mCherry/GFP signals, as depicted in [D]. Note that the experiment and statistics were performed together with the odc-1(RNAi) groups in Extended Data Fig. 6d . N = 121(CTL ad lib), 127(CTL fasted), 107( Dhps-1(RNAi) ad lib), 143( Dhps-1(RNAi) fasted) worms. (f) Representative fluorescence images of the head region of young C. elegans SQST-1::GFP fasted for two days and fed control or dhps-1(RNAi) expressing bacteria. Autophagic activity is indicated by a decrease in the number of GFP-positive particles. Scale bar = 50 μm. (g) Quantification of the SQST-1::GFP particles in the head region, as depicted in [F]. Note that the experiment and statistics were performed together with the odc-1(RNAi) groups in Extended Data Fig. 6f . N = 133(CTL ad lib), 86(CTL fasted), 108( dhps-1(RNAi) ad lib), 100( dhps-1(RNAi) fasted) worms. (h) Knockdown of dhps-1 does not affect worm size under ad lib or IF conditions. Note that the experiment and statistics were performed together with the odc-1(RNAi) groups in Extended Data Fig. 7g . N = 56(CTL ad lib), 66(CTL IF), 62( dhps-1(RNAi) ad lib), 59( dhps-1(RNAi) IF) worms. (i) Representative images of U2OS GFP-LC3 cells starved in HBSS with or without 100 µM GC7 for 6 hours. CQ was added for 3 hours before fixation. (j-k) Quantifications of [I]. N = 18 biologically independent samples. Statistics: [C,E,G] Kruskal-Wallis-test with Dunn’s correction. [H,J,K] Two-way ANOVA with Holm-Šídák’s multiple comparisons test. Bar graphs show the mean ± s.e.m. Source numerical data are available in source data.
Supplementary information
Supplementary information.
Supplementary Figs. 1–6 and uncropped immunoblots for Supplementary Figs. 5 and 6.
Reporting Summary
Supplementary table 1.
Supplementary Tables 1–5.
Supplementary Data 2
Numerical Source Data Supplementary Table 3.
Supplementary Data 1
Numerical Source Data for Supplementary Fig. 1.
Numerical Source for Supplementary Data Fig. 2.
Supplementary Data 3
Numerical Source Data for Supplementary Fig. 3.
Supplementary Data 4
Numerical Source Data for Supplementary Fig. 4.
Supplementary Data 5
Numerical Source Data for Supplementary Fig. 5.
Supplementary Data 6
Numerical Source Data for Supplementary Fig. 6.
Source Data Fig. 1
Numerical Source Data.
Source Data Fig. 2
Source data fig. 3.
Uncropped western blots.
Source Data Fig. 4
Source data fig. 5, source data fig. 6, source data extended data fig. 1, source data extended data fig. 2, source data extended data fig. 3, source data extended data fig. 4, source data extended data fig. 5, source data extended data fig. 6, source data extended data fig. 7.
Numerical Source Data
Source Data Extended Data Fig. 8
Source data extended data fig. 9, source data extended data fig. 10, rights and permissions.
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/ .
Reprints and permissions
About this article
Cite this article.
Hofer, S.J., Daskalaki, I., Bergmann, M. et al. Spermidine is essential for fasting-mediated autophagy and longevity. Nat Cell Biol (2024). https://doi.org/10.1038/s41556-024-01468-x
Download citation
Received : 04 April 2024
Accepted : 02 July 2024
Published : 08 August 2024
DOI : https://doi.org/10.1038/s41556-024-01468-x
Share this article
Anyone you share the following link with will be able to read this content:
Sorry, a shareable link is not currently available for this article.
Provided by the Springer Nature SharedIt content-sharing initiative
Quick links
- Explore articles by subject
- Guide to authors
- Editorial policies
Sign up for the Nature Briefing: Translational Research newsletter — top stories in biotechnology, drug discovery and pharma.

IMAGES
VIDEO
COMMENTS
For instance, in the field of biology, the invention of technologies (e.g., lenses, microscopes) allowed for detailed observation and rich descriptions of cells (i.e., qualitative research) that led to the development of cell theory, the establishment of the field of cell biology, and quantitative research on cell structure, function, and ...
Differences in a Nutshell In the world of research, there are two general approaches to gathering and reporting information: qualitative and quantitative approaches. Qualitative research generates non-numerical data while quantitative research generates numerical data or information that can be converted into numbers.
Here, we propose an approach able to identify when systemic qualitative changes in biological systems happen, thus opening the possibility for therapeutic interventions before the occurrence of ...
Qualitative research gathers participants' experiences, perceptions, and behavior. It answers the hows and whys instead of how many or how much. It could be structured as a standalone study, purely relying on qualitative data, or part of mixed-methods research that combines qualitative and quantitative data.
While old-style conservation biology was dominated by biological approaches and almost exclusively quantitative in its analytical techniques, the new conservation science frequently uses qualitative approaches developed in the social sciences to gather information, or determine why and how decisions are made.
Aim The aim of this paper is to provide an overview of qualitative research methods, including hands-on information on how they can be used, reported and assessed. This article is intended for beginning qualitative researchers in the health sciences as well as experienced quantitative researchers who wish to broaden their understanding of qualitative research.
Qualitative research involves the studied use and collection of a variety of empirical materials - case study, personal experience, introspective, life story, interview, observational, historical, interactional, and visual texts - that describe routine and problematic moments and meanings in individuals' lives.
To summarise the findings of conceptual change intervention studies in biology education, this qualitative review and meta-analysis endeavour to answer the following research questions:
Observation in qualitative research "is one of the oldest and most fundamental research methods approaches. This approach involves collecting data using one's senses, especially looking and listening in a systematic and meaningful way" ( McKechnie, 2008, p. 573). Similarly, Adler and Adler (1994) characterized observations as the "fundamental base of all research methods" in the ...
Educational research often requires mixing different research methodologies to strengthen findings, better contextualize or explain results, or minimize the weaknesses of a single method. This article provides practical guidelines on how to conduct such research in biology education, with a focus on mixed-methods research (MMR) that uses both quantitative and qualitative inquiries ...
10 Department of Biology, Duke University, Durham, NC, USA. An emerging challenge in plant biology is to develop qualitative and quantitative measures to describe the appearance of plants through the integration of mathematics and biology. A major hurdle in developing these metrics is finding common terminology across fields.
The flourishing plant science promotes the exploding number of data and the expansion of toolkits. Plant synthetic biology is still in its early stages and requires more quantitative and predictable...
Get definitions and examples of qualitative data and quantitative data. Learn how to tell the two types of data apart and when to use them.
Qualitative research methods treat non-numeric empirical materials as data. In other words, Qualitative researchers use words, images, sounds, video, and other data that have not been reduced to numbers to understand some research question of interest. This chapter explores what qualitative research entails and why researchers might want to ...
Author Summary We developed a process to quantitatively fit mathematical models using qualitative data, and applied it in the study of how stem cells are regulated in the fruit fly ovary. The available published data we collected are fluorescent images of protein and mRNA expression from genetic experiments. Despite lacking quantitative data, the new process makes available quantitative model ...
Qualitative research is a type of research that explores and provides deeper insights into real-world problems. Instead of collecting numerical data points or intervening or introducing treatments just like in quantitative research, qualitative research helps generate hypothenar to further investigate and understand quantitative data.
Indeed, a qualitative study by Watkins and Elby (2013) focusing on one student's interview about her views on mathematics in biology found that she held diverse, contextual views about the role of equations in understanding biology. Qualitative studies in K-12 have made important contributions to our understanding of biology epistemology.
Qualitative data in medical diagnosis involves the subjective assessment of symptoms, patient narratives, and contextual information. Some diagnosis tests are also qualitative or a hybrid of the two methods (semi-quantitative). Quantitative research is used to test hypotheses, establish relationships, and make predictions.
Qualitative research involves collecting and analyzing non-numerical data (e.g., text, video, or audio) to understand concepts, opinions, or experiences. It can be used to gather in-depth insights into a problem or generate new ideas for research. Qualitative research is the opposite of quantitative research, which involves collecting and ...
The scientific method conducts empirical research in such a way that it is without bias, is repeatable, and withstands the scrutiny of the scientific community. The opposite of the scientific method would be knowledge gained by testimonials. A testimonial would be where observations are made under non-controlled conditions.
Quantitative research in psychology and social sciences answers "how much" questions. Qualitative research answers the "how" and "why" of a phenomenon. Learn more.
Biology Research Topics Are you in need of captivating and achievable research topics within the field of biology? Your quest for the best biology topics ends right here as this article furnishes you with 100 distinctive and original concepts for biology research, laying the groundwork for your research endeavor.
This guide explains the focus, rigor, and relevance of qualitative research, highlighting its role in dissecting complex social phenomena and providing in-depth, human-centered insights.
Research article First published online August 7, 2024 The Experience of Phenylketonuria in Pregnancy and the Developing Maternal-Infant Relationship: A Qualitative Study
Virtual interviews have gained prominence among researchers from various fields as an effective method for qualitative data collection. The Covid-19 pandemic - with its movement restrictions,...
Hofer et al. show that fasting promotes the synthesis of spermidine, which stimulates eIF5A hypusination to induce autophagy and increase lifespan in various species in a conserved manner.
122 Remote Biology jobs available in Remote 🇮🇱 on Indeed.com. Apply to Biostatistician, Director, Research Analyst and more!